
Projected impacts of climate change on the habitat of Xerophyta species in Africa
2024-03-09 10:19VincntOkloWangaBoniacNgargaMillicntAkinyiOuloElijahMbandiMkalaVronicahMutlNgumbauGuyEricOnjalalainaWycliOchingOdagoConsolataNanjalaClintonOnyangoOchingMossKirgaGichuaRobrtWahitiGituruGuangWanHu
植物多樣性 2024年1期
Vincnt Oklo Wanga ,Boniac K.Ngarga ,Millicnt Akinyi Oulo ,Elijah Mbandi Mkala ,Vronicah Mutl Ngumbau ,Guy Eric Onjalalaina ,Wycli Oching Odago ,Consolata Nanjala ,Clinton Onyango Oching ,Moss Kirga Gichua ,Robrt Wahiti Gituru ,Guang-Wan Hu ,*
a CAS Key Laboratory of Plant Germplasm Enhancement and Specialty Agriculture,Wuhan Botanical Garden,Chinese Academy of Sciences,Wuhan 430074,China
b University of Chinese Academy of Sciences,Beijing 100049,China
c Sino-Africa Joint Research Center,Chinese Academy of Sciences,Wuhan 430074,China
d Centre for Integrative Conservation,Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Menglun 666303,China
e Botany Department,Jomo Kenyatta University of Agriculture and Technology,Nairobi,Kenya
f East African Herbarium,National Museums of Kenya,P.O.Box 451660-0100,Nairobi,Kenya
Keywords: Africa Climate change MaxEnt model Potential suitable distribution Velloziaceae Xerophyta
ABSTRACT Climate change poses a serious long-term threat to biodiversity.To effectively reduce biodiversity loss,conservationists need to have a thorough understanding of the preferred habitats of species and the variables that affect their distribution.Therefore,predicting the impact of climate change on speciesappropriate habitats may help mitigate the potential threats to biodiversity distribution.Xerophyta,a monocotyledonous genus of the family Velloziaceae is native to mainland Africa,Madagascar,and the Arabian Peninsula.The key drivers of Xerophyta habitat distribution and preference are unknown.Using 308 species occurrence data and eight environmental variables,the MaxEnt model was used to determine the potential distribution of six Xerophyta species in Africa under past,current and future climate change scenarios.The results showed that the models had a good predictive ability (Area Under the Curve and True Skill Statistics values for all SDMs were more than 0.902),indicating high accuracy in forecasting the potential geographic distribution of Xerophyta species.The main bioclimatic variables that impacted potential distributions of most Xerophyta species were mean temperature of the driest quarter(Bio9)and precipitation of the warmest quarter(Bio18).According to our models,tropical Africa has zones of moderate and high suitability for Xerophyta taxa,which is consistent with the majority of documented species localities.The habitat suitability of the existing range of the Xerophyta species varied based on the climate scenario,with most species experiencing a range loss greater than the range gain regardless of the climate scenario.The projected spatiotemporal patterns of Xerophyta species help guide recommendations for conservation efforts.
1.Introduction
The potential effects of climate change on biodiversity have received considerable attention.This is because climate change is expected to alter the growth and natural distribution of species,as well as increase biodiversity loss in natural habitats,resulting in a loss of species diversity and germplasm resources (Wan et al.,2021; Ngarega et al.,2021,2022a).By the year 2050,Africa is expected to experience a 2.6-3.0°C increase in average temperatures,and large areas of northern and southern Africa are projected to be >3.5°C above pre-industrial levels (Serdeczny et al.,2017;Lee et al.,2020;Mkala et al.,2022).It is projected that with the continued increase in global average temperatures and precipitation,Africa remains extremely vulnerable to climate change(Sintayehu,2018).
Climate variables influence the composition and distribution of vegetation,which can be a direct reflection of global climate change(Yan et al.,2021).Accordingly,researchers are focused on the linkages between biodiversity,ecosystem services,and climate change.These biological-climate predictions can serve as a foundation for hypothetical studies on species origin,vegetation partitioning,and floristic creation,as well as play a key role in biotechnology and species introduction and domestication (Zhang et al.,2020).However,most developing countries lack a comprehensive biodiversity inventory,as well as a sufficient grasp of particular species to identify those in danger (Burke,2004; ?oban et al.,2020).The classification of large geographic areas into biomes allows for an evaluation of vegetation types based on the principal climatic restrictions and related prevalent growth patterns(Burke,2004; Dar et al.,2020).
Climate change is estimated to be a major factor in the extinction of many animal and plant species (Nzei et al.,2021; Mkala et al.,2022).Species near a biome boundary and at the edge of their distribution ranges are likely to be the first to respond to climate changes (Thomas,2010).Thus,we can effectively target conservation initiatives to offset the effects of climate change by modeling their distributions (Merow and Jr,2014).Species distribution models (SDMs) are important for analyzing the spatial and temporal processes that make up a species’life cycle.Additionally,SDMs for historical timelines can also offer ecological and evolutionary data on the historical changes of species distribution over time (Park et al.,2022).Past models can be used to explain phylogeographic patterns and speciation processes,as well as to predict historical hotspots and potential migration routes (Park et al.,2022).Additionally,studies on how species have adapted to past climate change offer important insights into how species will respond to climate change in the future (Pearson,2006).Future distribution models can forecast habitat suitability and provide information on the likelihood of range shifts or population changes(Sinclair et al.,2010;Ngarega et al.,2022b).However,for the great majority of species,occurrence data is generally limited,especially for uncommon or extremely rare species (Merow and Jr,2014).MaxEnt is a popular tool for predicting the distribution of species based solely on occurrence point information(Merow et al.,2013).In addition,it can also predict habitat suitability for species in different habitats based on presence-only data (Elith et al.,2011;Gomes et al.,2018).Even though models do not precisely reflect the mechanisms that lead to observed distributions,they are useful in describing the relationships between species and environmental variables at wide spatial scales (Merow and Jr,2014).
XerophytaJuss.,is a monocotyledonous genus of the Velloziaceae family that is found on the African mainland,Madagascar,and the Arabian Peninsula(Beentje,1994;Elith et al.,2011;Farrant et al.,2015; Gomes et al.,2018).In mainland Africa,there are approximately 45 species,one species is found on the Arabian Peninsula,and 25 species are found in Madagascar (Behnke et al.,2013).The closest relative ofXerophytais the genusVelloziafound in South America,primarily Brazil(Mcpherson et al.,1997;Mello-Silva et al.,2005; Alcantara et al.,2015; Wanga et al.,2021).The family Velloziaceae has been shown to have a Gondwanan origin,with the separation between African and South American species corresponding to the continents dividing roughly 100 million years ago(Mello-Silva et al.,2011).All African species are desiccation tolerant,i.e.,resurrection plants(Alcantara et al.,2018).Most of theXerophytaspp.,exceptXerophyta elegans,are poikilochlorophyllous,i.e.,they lose their chlorophyll during desiccation,having protective mechanisms that are fully activated in the leaves(Mello-Silva et al.,2011;Behnke et al.,2013;Farrant et al.,2015).Most research on the genus has focused on the whole genome and the mechanisms that underlie desiccation tolerance in these species(Farrant et al.,2015;Costa et al.,2017; Radermacher et al.,2019; Wanga et al.,2020;2021),which can be key in tackling food security amidst the global warming threats.Recent reports indicating the high frequency of fire occurrence and human exploitation (i.e.,brooms or brushes),medicinal products and fuelwood have been recorded to impact the habitat ofXerophytaspecies,necessitating urgent conservation actions(Goodman,2021;IUCN,2023).To date,there have been no studies that have simulated or projected the distribution ofXerophytaspecies.This lack of attention may be attributed to the fact that these species grow in inaccessible areas,hence they are botanically unexploited(Behnke et al.,2013).As a result,examining the factors that impact the distribution and evolution ofXerophytain Africa could offer valuable insights into how xerophytic species respond to changes in climate.
In this study,we explored the potential distribution of sixXerophytaspecies under past,current and future climates using the MaxEnt model.We utilized two Representative Concentration Pathway (RCPs) scenarios representing moderate (RCP 4.5) and extreme greenhouse gas emissions(RCP 8.5)to model the potential distribution ofXerophytaspecies in the 2070s.The past climate was represented by the Last Glacial Maximum(LGM,22,000 ya)and the Mid-Holocene (MH,6000 ya).The goals of the study were to (1)understand the geographical distribution patterns and future changes ofXerophytaspp.in Africa,(2) determine the dominant bioclimatic variables that limit the habitat distribution ofXerophytaspp.,and (3) model suitable habitat ofXerophytaspp.under past,current and future climate change scenarios and provide a scientific basis for the protection ofXerophytaspp.resources.
2.Materials and methods
2.1.Species occurrence data
There areca.70Xerophytaspecies native to tropical Africa and Madagascar (Wanga et al.,2021).However,several species lack adequate distribution data to allow for successful modeling(Stockwell and Peterson,2002).In this study,six species with adequate and validated occurrences were studied.We obtained species occurrence points from the Global Biodiversity Information Facility (GBIF,http://www.gbif.org/; accessed in March 2021).Because some of these occurrences lacked geographical information,Google Earth was utilized to supplement the latitude and longitude information.Spatial thinning of occurrence data is a simple and easy-to-implement strategy for mitigating the impacts of sampling bias(Aiello-Lammens et al.,2015).We used thespThinpackage (Aiello-Lammens et al.,2015) in the R environment to ensure that only one occurrence record per grid cell at a resolution of 2.5 arc-min.After spatial thinning of occurrences,points obtained were used for modeling;69 forXerophyta dasylirioides,33 forX.elegans,63 forX.humilis,30 forX.pectinata,92 forX.viscosa,and 21 forX.villosa(Table S1; Fig.1).ArcGIS 10.5 (Esri,Redlands,CA,USA,http://www.esri.com) was then used to create maps of the remaining species occurrences (Fig.1).
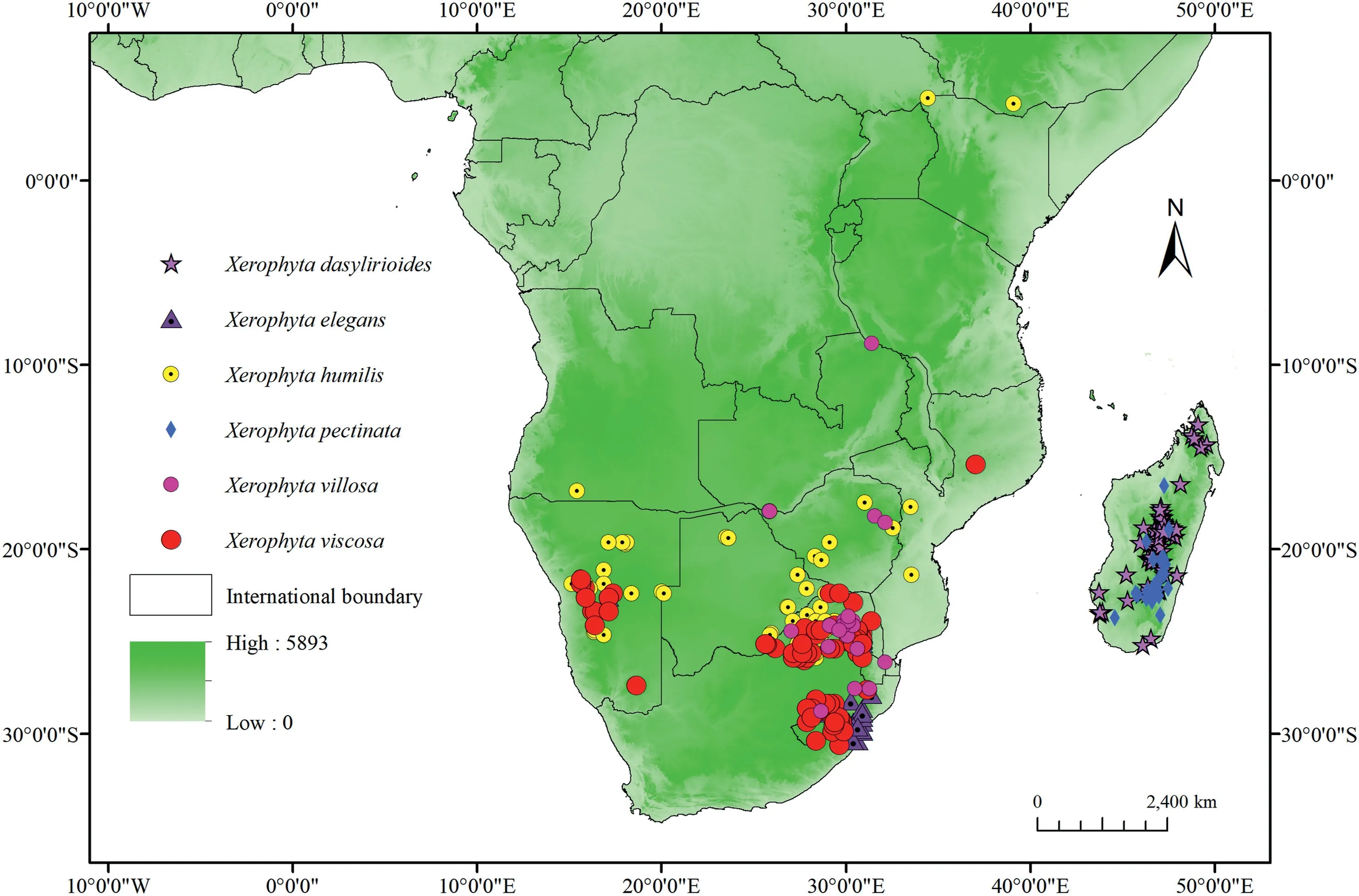
Fig.1.Locality and occurrence records of Xerophyta species employed in the modelling.
2.2.Environmental variables
We employed ‘bioclimatic’ characteristics from the World-Clim2.1 database (https://www.worldclim.org/data/worldclim21.-html) to characterize climatic conditions relevant to biological species(Table S2;Fick and Hijmans,2017).This data set is based on monthly consensus climatologies spanning 1970 to 2000.The data set was obtained at a spatial resolution of 2.5 arc-min or 5 km2,which roughly corresponds to the uncertainty inherent in the occurrence data.We removed nine data layers (described in Table S3) because they contained artifacts that would otherwise cause sharp variances and multicollinearity between neighboring pixels.We excluded the variables with correlation coefficients of more than 0.7.This analysis was achieved through the Variance Importance Factor(VIF)procedure in theusdmpackage available in R platform v.3.6.2(R Core Team,2022).Data for the past and future scenarios were obtained from WorldClim1.4 and WorldClim2.1,respectively,with a spatial resolution of 2.5 arc-min(Hijmans et al.,2005; Fick and Hijmans,2017).We utilized one global climate model (GCM),the Community Climate System Model version 4(CCSM4; Gent et al.,2011),to download the climate data for the future(2070s)and for the past.Climate data for the past came from 22,000 years ago,reflecting the LGM,and 6000 years ago,reflecting the MH.This GCM(CCSM)has been recognized as one of the most appropriate GCMs for the African region(Nzei et al.,2021;Ngarega et al.,2022a).The RCP scenarios consist of four pathways,including RCP 2.6,RCP 4.5,RCP 6.0,and RCP 8.5(Remya et al.,2015).RCP 4.5 and RCP 6.0 are both moderate greenhouse gas emission scenarios,with RCP 4.5 representing a scenario in which a higher priority is given to mitigation than in RCP 6.0 (Wei et al.,2018).RCP 8.5 implies that no climate policies would be implemented in the future or the present,and it reflects very high emissions scenarios(1370-ppm CO2equivalent by 2100; Van Vuuren et al.,2011).Therefore,we chose RCP 4.5 (moderate scenario) and RCP 8.5 (extreme scenario)to simulate the suitable habitat distribution ofXerophytaspp.for the future time frame.
2.3.Model calibration and development
We utilized MaxEnt 3.4.4 (Elith et al.,2011; Phillips and Dudík,2008) to predict the distribution ofXerophytaspecies.MaxEnt was firstly used to build a model,and then the predict function was used in conjunction with a specified set of independent variables to calculate the likelihood of occurrence.The potential distribution of our focal taxa was predicted using the climatic variables and occurrence localities described in the earlier steps.For model calibration and development,25% of the occurrence data were chosen at random as test sets and 75% as training sets.Following the ‘Subsample’ strategy,the model was duplicated ten times.The background region represents accessible space that can be colonized or has previously been inhabited by the species being modeled (Barve et al.,2011).Like prior studies,we used 10,000 random background points (Nzei et al.,2021).The output format was set to‘Logistic’,with a maximum iteration of 5000.The default settings for the rest of the parameters were kept.The area under the receiver operating characteristic(ROC)curve(AUC;Area under the ROC Curve) and True Skill Statistics (TSS) were used to assess the accuracy of the MaxEnt models (Allouche et al.,2006; Warren and Seifert,2011; Ngarega et al.,2022b).AUC values vary from 0 to 1,and the closer value is to 1,the better the model’s performance.The accuracy of the model is rated in five categories: Fail(0.5-0.6),Poor (0.6-0.7),Fair (0.7-0.8),Good (0.8-0.9),and Excellent(0.9-1.0),with a value less than 0.5 equaling randomness(Swets,1988).The jackknife test was also used to assess the impact of variables on theXerophytaspecies.Results of jackknife tests show the weight of different environmental factors affecting the habitat suitability ofXerophytaspecies.The final outputs were continuous suitability layers made up of the mean of the predictions from the ten model runs.To illustrate the differences and convergences between each model,the average continuous logistic output predictions were transformed into binary maps using the Maximum sum of training sensitivity and specificity (MTSS)threshold value that maximizes sensitivity and specificity(Table S4; Liu et al.,2016).
2.4.Changes in habitat suitability and niche overlap analysis
We used the Python-based GIS toolkit SDMtoolbox(Brown et al.,2017) in ArcGIS v.10.5,to investigate the differences between the current(baseline)and past or future climate distribution areas.We imported the binary maps of suitability described in the previous steps into ArcGIS v.10.5 and used the SDMtoolbox to analyze and map the spatial range changes of the suitable habitat area ofXerophytaspp.
The examination of niche overlap allowsXerophytaspecies to estimate the niche they share.The logistic output maps for the sixXerophytaspecies pairwise pairings were compared to analyze the niche overlap between the species using Schoener’sDmetrics(Warren et al.,2008).This analysis was implemented in ENM tools v.1.4.4 (Warren et al.,2010).With values ranging from 0 to 1,this metric assesses the niche overlap of two species,with 1 indicating similarities in the two niches and 0 indicating no overlap (Warren et al.,2008).
2.5.Multivariate environmental similarity surface (MESS) analysis
When creating models to determine how climate change affects species'suitable areas,a common challenge is understanding to what extent future climatic conditions differ from contemporary climatic variables used in the model.One method to measure this is by using multivariate environmental similarity surface(MESS),as outlined by Elith et al.(2010).The MESS method involves identifying a reference layer of environmental variables and calculating the similarity (S) between the point set of environmental variables in the reference layer and those under different climatic conditions.A positive S value indicates a climate difference at the point,with a smaller S value indicating a more significant difference.Conversely a negative S value shows that at least one environmental variable at that point is outside the reference range,indicating a considerable environmental change.We applied the MESS procedure by utilizing the ‘density.-tools.novel’ tool in the maxent.jar file and plotted the results for the past and future climate scenarios.
3.Results
3.1.Performance of the model and variable contribution
After correlation analyses,the remaining variables included the mean diurnal range(Bio2),isothermality(Bio3),mean temperature of wettest quarter (Bio8),mean temperature of driest quarter(Bio9),precipitation of wettest month (Bio13),precipitation of driest month(Bio14),precipitation of warmest quarter(Bio18),and precipitation of driest quarter (Bio19) (Tables S2 and S3).The modeling of the current distribution ofXerophytaspecies yielded strong evaluation scores for AUC and TSS values greater than 0.902,indicating that the MaxEnt model was very effective in forecasting the potential distribution for theXerophytaspecies (Table 1).Jackknife results indicated that when used in isolation,precipitation of warmest quarter (Bio18) influenced the distribution ofX.dasyliroidesandX.pectinatamost,indicating that this variable contains considerable information absent in other variables (Fig.2).The impact of bioclimatic conditions on species distribution was assessed using the percent contribution (Table 2).The minimumtemperature of the coldest quarter (Bio9) and precipitation of the warmest quarter (Bio18) were the principal bioclimatic variables impacting the potential distribution of the majority of the species.For all the studied species,the precipitation of the coldest quarter(Bio19) and the precipitation of the warmest quarter (Bio8) had a low contribution.This implies that the minimum temperature and precipitation of the warmest quarter are the most critical factors influencing theXerophytaspecies’ potential distribution.On the other hand,jackknife analyses showed that when used in isolation,isothermality(Bio3),precipitation of warmest quarter(Bio18),and mean temperature of driest quarter (Bio9) had the most gains for different species (Fig.2).

Table 1 Model performance evaluation for six Xerophyta species using Area Under the Curve(AUC) and True Skill Statistics (TSS) analyzed in MaxEnt.
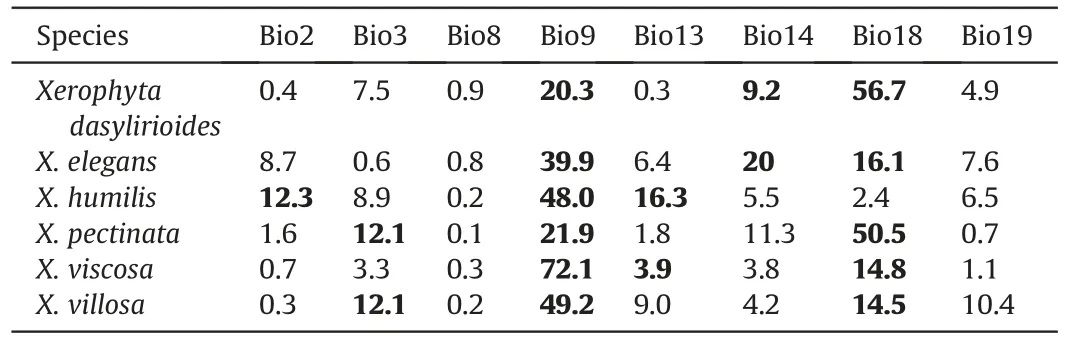
Table 2 Variable contribution to the final MaxEnt models of Xerophyta species.
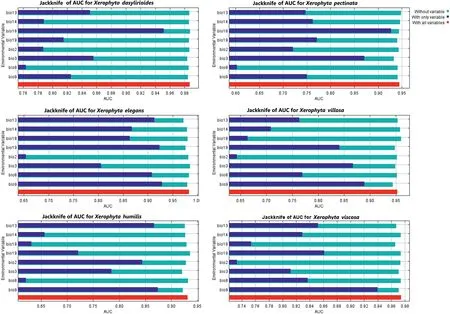
Fig.2.Jackknife results for Xerophyta spp.distribution using MaxEnt models.
3.2.Predicted potential distribution of Xerophyta
The potential distribution ofXerophytain the past (LGM and MH),present,and future(RCP 4.5 and RCP 8.5),are shown in Fig.3.Suitability regions were classified into high,medium (moderate),low and no species suitability regions.Environmental conditions during the LGM were less suitable forXerophytaspecies than were those of the MH.However,environmental conditions between these two past periods did not significantly differ forX.pectinata,X.humilisandX.villosa.This allowed the geographic expansion,reduction and even the stable regions for the differentXerophytaspecies under current climatic conditions.
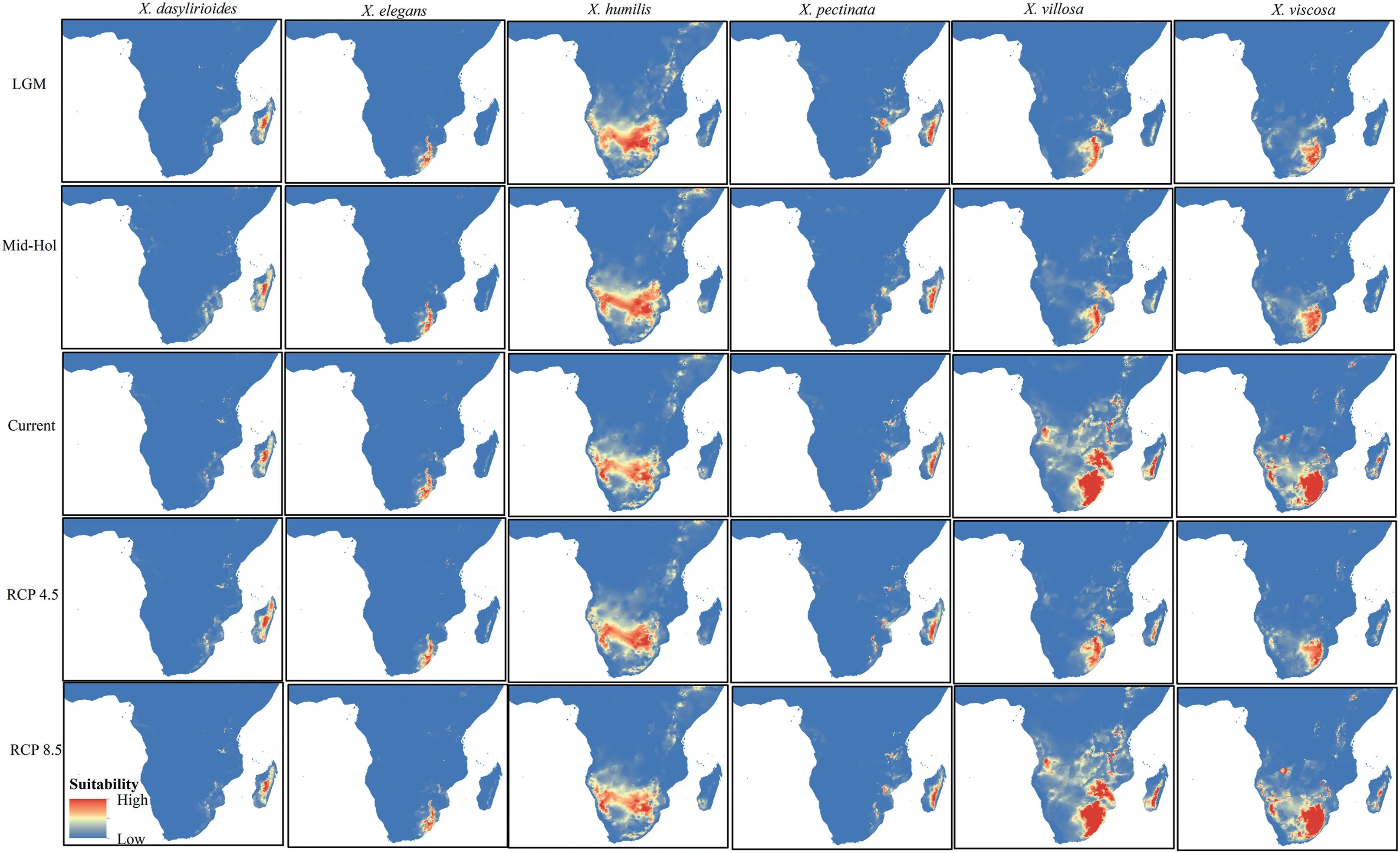
Fig.3.Xerophyta spp.prediction maps for the Last Glacial Maximum (LGM),Mid-Holocene (Mid-Hol),current climatic conditions,future,RCPs 4.5 and 8.5.Species distribution in their habitats are color coded differently to show high,medium and low suitability areas.Red indicates high suitability,brown indicates medium suitability,white indicates low suitability and blue indicates no species distributed in the area.Habitat suitability ranges from 0 (blue,low suitability) to 1 (red,high suitability).
Baseline current models revealed that the moderate and highsuitability zones forXerophytaare mostly found in tropical Africa and correspond to the recorded occurrence localities.X.humilis,X.villosa,andX.viscosawere projected to have a continuous highsuitability distribution,whereas that ofX.pectinatawas fragmented.The potential distribution for these taxa varied across the region.For instance,X.villosawas projected to have potential distribution areas in Zambia,Zimbabwe and South Africa,whereasX.humiliswas projected to have potential distributions in Angola,Botswana,Ethiopia,Mozambique,Namibia,Northern Provinces,Sudan,Swaziland,Zambia,and Zimbabwe (Fig.3).The species’potential current suitable habitats forXerophytaranged from 166.05 × 103km2(X.elegans) to 1915.75 × 103km2(X.humilis)(Table S5).
3.3.Patterns of change in habitat suitability
To clarify how geographic distribution patterns ofXerophytaspecies may change and identify predicted migration paths,we calculated the change in suitability compared to current suitability for each grid cell and scenario.These useful changes are shown in Fig.3,which show areas where contraction(decrease in fitness over time)and expansion(increase in fitness over time)occur.The range possibilities for suitability changes cover four different situations:large expansions from low suitability,large contraction from high suitability,constant high suitability,and constant low suitability.Range change differed between past and the two future scenarios depending on each species’ response.For most species,range loss was predicted to be larger than range gain,irrespective of the scenario(Fig.4),however,the greatest losses were predicted under the RCP 8.5 scenario (Table S5).Specifically,under RCP 8.5,X.pectinata(35.40%),was predicted to lose the largest portions of its suitable ranges,whereasX.dasylirioides(129.1%) was predicted to have the highest gains in suitability.

Fig.4.Representation of the past and future distribution maps of Xerophyta species range change for Last Glacial Maximum (LGM),20,000 ya,Mid-Holocene (Mid-Hol),6000 ya,RCP 4.5 and RCP 8.5.Stable areas are shown in purple,areas of loss are shown in red,areas of gain are shown in green,and unsuitable areas are shown in grey.
3.4.Niche comparison
According to Schoener’sDmetrics,the niches ofXerophytaspecies overlap considerably (Table 3).The highest niche overlap was predicted inX.pectinataandX.dasylirioides(0.635).Other species,such asX.humilisandX.pectinata,however,are projected to occupy different ecological niches.

Table 3 Comparisons of ecological niche overlap for Xerophyta species using the Schoener’s D statistics.
3.5.Multivariate environmental similarity surface analysis of Xerophyta potential distribution areas under different climate scenarios
The region where the climate anomaly(S <0)could potentially occur was small in reference to the potential distribution area of allXerophytaspecies,as shown in Fig.5,under the past and future climate scenarios.Notably,compared to all other climate scenarios for all theXerophytaspecies,the RCP 8.5 was observed to have the largest degree of extrapolation(Fig.4),indicating that this scenario had the highest degree of anomaly.

Fig.5.The multivariate environmental similarity surface (MESS) map by data source (Xerophyta spp.localities) and considering the environmental variables as reference environmental area for the African region for the past (LGM and Mid-Hol) and future distribution (RCPs 4.5 and 8.5).
4.Discussion
Niche modeling is a useful tool for predicting the potential impacts of climate change on the distribution of species as well as their responses (Ngarega et al.,2022b).This is the first study to use a niche modeling approach to model the distribution ofXerophytain Africa.The resulting outputs of our study may provide valuable information for conservation,management and research of the genus.Here,we specifically utilized the MaxEnt modeling approach to project areas that have suitable climates for sixXerophytaspecies under the current baseline period and evaluated the impacts of climate change on their past and future potential habitat distribution.The MaxEnt models had very high predictive capacities (AUC and TSS values above 0.902; Table 1),which is comparable to various studies done in the region using MaxEnt modeling approach (Nzei et al.,2021; Wan et al.,2021),as well as other studies beyond the African region (Yan et al.,2021).
4.1.Significant variables affecting Xerophyta distribution
The potential distribution of plant species is generally affected by climate variables,which contribute significantly to their growth and abundance(Beamount et al.,2005).We evaluated the key bioclimatic variables determining the variance inXerophytaspecies distribution based on the results of the jackknife test and the contribution rates obtained by the MaxEnt model (Wiens et al.,2009; Zhu et al.,2013).Our results indicate that the distributions ofXerophytaspecies are mainly affected by the following variables:(1)minimum temperature of the coldest quarter(Bio9),and (2) precipitation of the warmest quarter (Bio18) (Table 2).These results indicate that both temperature and precipitation variables limitXerophytadistribution.These findings areconsistent with a previous study that showed freely drained sandy or rocky soils,high temperatures and seasonal rain regimes favor the establishment of resurrection and drought-resistant plants(Alcantara,2018).
Species of the genusXerophytaare well-adapted to xeric environments and all tested species display vegetative desiccation tolerance with most of the species being poikilochlorophyllous and others homoichlorophyllous (Lyall et al.,2020).This may explain the variations that occurs in the range change responses of the species,with most species having a restriction to the inselbergs during extreme conditions hence forming terrestrial islands for survival(Chen et al.,2019).Recent studies have shown that most species ofXerophytaexhibit a series of changes in their physiologies when exposed to high temperatures,including chlorophyll degradation,cessation of growth,and senescence,an expression of desiccation tolerance (Costa et al.,2017;Radermacher et al.,2019).Moreover,some species have adapted to these conditions,e.g.,X.viscosawas shown to express the geneXVSAP1at both high and low temperatures,which conferred higher adaptability to these hostile environments (Garwe et al.,2003).Finally,the harsh chasmophytic habitat of most of its species leads to frequent periods of severe water deficit,even during the wet seasons (Farrant et al.,2015).Therefore,in high temperature and high precipitation areas of Africa,the distribution patterns of theXerophytaspecies are predicted to frequently shift in the future.
4.2.Ecological niche suitability projections for the Last Glacial Maximum and Mid-Holocene
The habitat suitability projected for the LGM and MH scenarios differed from those obtained for current climate conditions.The habitat suitability projected for the LGM and MH scenarios varied among theXerophytaspecies with most species preferring LGM environmental conditions for persistence.According to the LGM projections,the species occurred in the tropical Africa,Southern and Central Africa,and the Madagascar regions,however,they did not differ much with the MH period,except for a reduction in range size.These regions were considered suitable putative cryptic refugia that persisted during the LGM.Refugia are areas in which a population can survive through an extended period of unfavorable conditions,and have hence facilitated the persistence of components of biodiversity over long periods of time and through changing climatic conditions (Keppel et al.,2011).Notably,when environmental conditions turn out to be more suitable for the species,they could potentially expand from within these refugia.During the LGM,the climate was cooler and drier,hence no rainfall;thus,vegetation only occurred in high-elevation refugia regions due to the formation of mist(Leal,2004).However,during the MH the temperature became warmer (Park et al.,2019),hence those species that were widely adapted could spread.Additionally,the difference between the two climate scenarios of the past could be a result of desiccation tolerance speciation,evolution and adaptation for theXerophytaspecies,which would explain the differences in the species responses among the regions (Lyall et al.,2020).
4.3.Implications of future climate scenarios to species distribution and conservation
Climate change affects species distribution,and future projections suggest that it will become more intense,leading to global warming (Putten et al.,2010).In addition,the impacts of climate change depend on various combinations of traits in plant species,which determine their abundance,resistance to disturbance and ability to respond to changes in environmental conditions.Our results showed that the current suitable habitat forXerophytaspp.ranges from 166.05 × 103km2(X.elegans) to 1915.75 × 103km2(X.humilis) (Table S5).In addition,models showed that highly suitable habitats are in the tropical regions,which are concurrent with recorded species localities.The prediction models also indicate that highly suitable habitat is currently available in Eastern,Central and Southern Africa for most of theXerophytaspecies.
Our models predicted that habitat loss,gain,and stability would vary in the future for differentXerophytaspecies.For instance,X.pectinata,andX.humiliswere projected to lose the largest portions of their suitable ranges,whereasX.dasylirioideshad the greatest range gains (Table S5; Fig.4).The varying responses under the high concentration emission scenario,RCP 8.5,could be due to the poikilochlorophyllous and homoiochlorophyllous nature of species in this genus.The results clearly show that predicted future climatic changes will have a significant impact on the geographic distribution ofXerophyta; the ability of plants in this genus to adapt or move to new locations with more favorable environmental conditions will be critical for the survival of the genus in the future.As a result,desiccation tolerance mechanisms may play important roles in the future distribution of the species,thereby warranting further investigations.It is also possible thatXerophytawill not adapt to new developments,and the loss of their habitat range,as predicted in this study,may be a representation of the disappearance of geographic loss.It is important to note that climate change effects on the genus might differ among individual species owing to differences in genetic and physiological tolerance of the different species.SomeXerophytaspecies share similar habitats and niches since their ranges overlap.Considering the significance of the mean temperature of the driest quarter (Bio9) and precipitation of warmest quarter(Bio18) in determining the habitat suitability ofX.pictinataandX.dasylirioides,it is reasonable to observe a high degree of overlap in their niches as demonstrated in Table 3.
The current study evaluated the potential impacts of past and future climate change on the distribution of sixXerophytaspecies in Africa,thereby,providing basic information on this little-known genus.The high level of agreement between the predicted distributions and the botanical collections revealed that the climatic variables studied are certainly crucial in determiningXerophytadistribution patterns.However,the accuracy of our MaxEnt model predictions may be limited by various factors,including the methodology and data applied herein.For example,our data were mostly gathered from GBIF and online databases,corresponding to low occurrence points for several species,which may have resulted in biases in the modeling approach.Secondly,MaxEnt modelling may not account for other factors impacting local adaptation,dispersal,and lack of precise knowledge about changes in land use throughout the predicted range ofXerophytaspp.Due to the paucity of research in these inselbergs where theXerophytaspecies occurs,we were unable to identify any of the additional variables impacting distribution,adaptability,and evolution,which should be considered in future research.As a result,while the results offer insights into the distribution ofXerophyta,the model outputs of our study should be treated with caution.
5.Conclusion
In the present study,we modeled the past and current distribution of suitable habitats for sixXerophytaspp.and evaluated how climate change would affect their distribution in the future using the Maxent algorithm.Our findings reveal that the habitats suitability among theXerophytaspecies are predicted to vary owing to climate change.Nonetheless,with the changing climate,the suitable habitat ofXerophytaspecies may be compromised.In addition,with the low records and poor sampling ofXerophyaspecies in Africa,aggressive species documentation and taxon sampling may be required to enhance our knowledge of the occurrence extent and area of occupancy for the genus.Finally,additional studies combining ecology,phylogeography and behavior are required to improve the understanding of the ecology and evolution ofXerophytain Africa.
Data availability statement
The data sets presented in this study can be found in the online repositories(GBIF) and supplementary materials.
Author contributions
VW,BN,VN,GWH,RG:conceptualization.GWH and RG:funding acquisition.GWH,MG and RG: supervision.VW and BN: methodology.VW,BN,MO,EM,CN,CO,GO and WO.VW,BN,GWH,MG,MO,CO and EM revised the draft manuscript.All authors have read and agreed to the final version of the manuscript.
Declaration of competing interest
The authors declare no conflict of interest.
Acknowledgements
We are sincerely thankful to the anonymous reviewers for comments and recommendations that helped improve this manuscript.This work was supported by grants from the International Partnership Program of Chinese Academy of Sciences(151853KYSB20190027),Sino-Africa Joint Research Center,CAS(SAJC202101) and The ANSO Scholarship for Young Talents,PhD Fellowship Program University of Chinese Academy of Sciences,China.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2023.05.001.

- 植物多樣性的其它文章
- Flower morphology of Allium (Amaryllidaceae) and its systematic significance
- Phylotranscriptomic discordance is best explained by incomplete lineage sorting within Allium subgenus Cyathophora and thus hemiplasy accounts for interspecific trait transition
- How to fill the biodiversity data gap:Is it better to invest in fieldwork or curation?
- Life forms affect beta-diversity patterns of larch forests in China
- A phylogenetic approach identifies patterns of beta diversity and floristic subregions of the Qinghai-Tibet Plateau
- Community structure and species diversity dynamics of a subtropical evergreen broad-leaved forest in China: 2005 to 2020