
Megafossils of Betulaceae from the Oligocene of Qaidam Basin and their paleoenvironmental and phytogeographic implications
2024-03-09 10:19ToYngJiHoCiYnZhiDiHongYuChenLeiHnLiZhngWeiYuLingXuJunLiWenJiLiJingYuWuSnPingXieDeFeiYn
植物多樣性 2024年1期
To Yng ,Ji-Ho Ci ,Yn-Zhi Di ,Hong-Yu Chen ,c,Lei Hn ,Li Zhng ,Wei-Yu Ling ,Xu-Jun Li ,Wen-Ji Li ,Jing-Yu Wu ,Sn-Ping Xie ,De-Fei Yn ,c,*
a School of Earth Sciences and Key Laboratory of Mineral Resources in Western China (Gansu Province),Lanzhou University,Lanzhou 730000,China
b Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences,Beijing 100044,China
c State Key Laboratory of Palaeobiology and Stratigraphy,Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment,Chinese Academy of Sciences,Nanjing 210008,China
d Center for Research and Education on Biological Evolution and Environments,School of Earth Sciences and Engineering,Nanjing University,Nanjing 210023,China
Keywords: Paleoenvironment Biogeography Betulaceous fossil Qaidam basin Tibetan Plateau Oligocene
ABSTRACT Understanding the paleoenvironment and phytogeographical history of the Tibetan Plateau,China relies on discovering new plant fossils.The Qaidam Basin has long been regarded as an ideal ‘field laboratory’to investigate the paleoclimate and paleobiological evolution of the northern Tibetan Plateau.However,fossil angiosperms from the Qaidam Basin are rare,and our knowledge of its paleovegetation is poor.Here,we report fossil leaves and fruits of Betulaceae found from the Oligocene Shangganchaigou Formation of northwestern Qaidam Basin (Huatugou area).Comparative morphological analysis led us to assign the fruits to the Betula subgenus Betula and the leaves to Carpinus grandis.These findings,together with other reported fossil plants from the same locality,reveal a close floristic linkage between the Qaidam Basin and Europe during the Oligocene.The northern pathway of this floristic exchange may have crossed through the Qaidam Basin during the late Paleogene.This floristic linkage may have been facilitated by the continuous narrowing of the Turgai Strait and stronger westerlies,which transported moisture and provided favorable climatic conditions.Indeed,fossil plants collected from the Qaidam Basin suggest that during the Oligocene this region had warm and humid deciduous broad-leaf forest,which differs from the region’s modern vegetation and indicates that the Qaidam Basin may have been a suitable region for these plants to flourish and spread during the Oligocene.
1.Introduction
The Tibetan Plateau,known as‘the roof of the world’,is an ideal natural laboratory for studying biological evolution in the Cenozoic(Chang and Miao,2016).The biogeographic history of the Tibetan Plateau,especially regarding its connections with other regions in the Northern Hemisphere during the Cenozoic,is quite complex.Researchers have made explored the molecular phylogenetic and biogeographic history of the Tibetan Plateau based on modern species from this region(Renner,2016;Ding et al.,2020).In recent years,several hypotheses have been proposed on the biogeographic significance of the Tibetan Plateau: Into Tibet,Out of Tibet,Out of India,and Into and Out of Africa(Deng et al.,2011;Wen et al.,2014;Zhou et al.,2022).Most of these were proposed to explain the biogeographical connections between the Tibetan Plateau and other regions during the Neogene or Quaternary.A series of excellent works in recent years have revealed high terrestrial biodiversity on the Tibetan Plateau during the Paleogene(Sun et al.,2015;Wu et al.,2017;Ai et al.,2018;Xu et al.,2018;Su et al.,2018,2019,2020; Deng et al.,2019a,2019b,2020; Liu et al.,2019; Low et al.,2019; Tang et al.,2019; Huang et al.,2020).Recently,fossil assemblages from the middle Eocene of central Tibet indicated a relatively free floristic exchange across the Northern Hemisphere(Su et al.,2020).Fossil plants collected from different strata in Tibet indicate extensive biogeographical connections with other floras of the Northern Hemisphere and make Tibet a crossroad for global floristic exchange in the Paleogene(Zhou et al.,2022).It is crucial to reconstruct the pathways for the floristic linkage between the Tibetan Plateau and other Northern Hemisphere regions during the Paleogene (Tang et al.,2019).
The Qaidam Basin is the largest Cenozoic terrestrial intermountain basin on the Tibetan Plateau (Fang et al.,2007; Miao et al.,2016).It has been regarded as a key region to study the paleoclimatic and paleobiological evolution of the northern Tibetan Plateau(Wang et al.,2007).Angiosperms play an important role in global ecosystems today; meanwhile,fossil angiosperms can provide important information on Cenozoic climate change(Sun et al.,2002).However,compared with abundant Cenozoic vertebrate fossils found in the basin (Wang et al.,2007,2015; Chang et al.,2008; Yang et al.,2018,2022),the fossil record of angiosperms from the Paleogene Qaidam Basin is relatively small.The plant records published thus far include Simaroubaceae Candolle,Leguminosae Jussieu,Salicaceae Mirb.,and Juglandaceae DC.ex Perleb,and are mainly from the Huatugou area and Dahonggou area,which are located in the northwestern and northern margins of the basin,respectively (Yan et al.,2018; Han et al.,2020; Song et al.,2020;Yang et al.,2021a,2021b).Information on the Paleogene paleovegetation in the basin is still incomplete.Nevertheless,preliminary work has indicated that in the late Paleogene the vegetation of this region was very different from that of modern times.
Betulaceae Gray,which consists of six genera (CarpinusL.,CorylusAscherson,OstryaScop.,OstryopsisDecne,AlnusMill.,andBetulaL.) and 150-200 species,is an important component of the deciduous broad-leaf forest in the north temperate zone today (Li and Skvortsov,1999).Betulaceous fossils are common in Cenozoic floras of the Northern Hemisphere (Crane,1981; Chen,1994a,1994b; Liu,1996; Lin et al.,2010; Liu et al.,2014; Xue et al.,2020).They are often found in different floras such as northern temperate deciduous broad-leaf forest,subtropical deciduous broad-leaf forest,and subtropical evergreen and deciduous broad-leaf mixed forest (Tao and Du,1987).Betulaceous fossils not only provide information about paleovegetation,but also contribute to reconstructing Cenozoic paleobiogeograhic and paleoclimatic history(Chen,1994a,1994b).Herein,fossil leaves and fruits of Betulaceae from the Oligocene Shangganchaigou Formation of northwestern Qaidam Basin,includingBetulaandCarpinus,are described from the Oligocene Shangganchaigou Formation of northwestern Qaidam Basin.The fossil fruits are assigned toBetulasubgenusBetula,and the fossil leaves are attributed toCarpinus grandisUnger emend.Heer.based on morphological comparisons.These fossils,together with other reported fossil plants from the same locality,not only enrich our knowledge of the paleovegetation in the Qaidam Basin,but also provide insights into the intercontinental floristic linkage between the Tibetan Plateau and other regions of the Northern Hemisphere during the Oligocene.
2.Materials and methods
The new specimens were collected from the Huatugou region(N38°24ˊ41ˊˊ,E90°53ˊ14ˊˊ,and N38°24ˊ38ˊˊ,E90°53ˊ9ˊˊ),the middle portion of the Shangganchaigou Formation,Qinghai Province,NW China at an elevation of about 3500 m a.s.l.All of the fossil leaves were unearthed from the same locality in the same layer,whereas the fossil fruits were collected from a different layer that is stratigraphically ~100 m lower (Fig.1).The formation,which was deposited in a lacustrine environment,is lithologically composed of conglomerates,sandstones,siltstones,mudstones,micritic carbonates,and gypsum beds.Our fossils were collected from lightgreen and light-yellow silty mudstone.Recent magnetostratigraphic and paleobotanic studies have dated the Shangganchaigou Formation to the Oligocene (Sun et al.,2005; Lu and Xiong,2009;Chang et al.,2015;Ji et al.,2017;Song et al.,2020).According to the‘interpolation method’ (He et al.,2021),if the lithofacies and deposition rates are assumed to be uniform,then the bottom and top ages of the formation can be used as primary markers,and the thickness relative to the top or base of the unit can be used to interpolate the ages.The lithofacies and deposition rate of Shangganchaigou Formation in the Huatugou Section,according to detailed study (Chang et al.,2015) and our observation in the geological survey,are uniform;so,we estimate that the age of our betulaceous fossils is about 28-26 Ma (early Chattian) based on their position in the section.

Fig.1.(a) Map showing detailed location of the fossil site in the Qaidam Basin,Qinghai Province,China.(b) Stratigraphic column of the Oligocene Shangganchaigou Formation.(c-d) The fossil-bearing layer in sites HTG022 and HTG028.(e) Living Carpinus betulus about 120 years old in the arboretum Kornik,Poland (from Boratynski,2014).(f) Local topographic features surrounding the present fossil site,Huatugou (approximately 3500 m a.s.l.).
All the specimens described in this study were observed under a Leica MZ 12.5 stereo microscope and figures were prepared using Adobe Photoshop CC 2015.Specimens ofBetulaandCarpinusused for comparison were retrieved from the Trinity Herbarium,Trinity College(Dublin,Ireland),Xishuangbanna Tropical Botanical Garden(Mengla,China),and Herbarium of Lanzhou University (Lanzhou,China).Cleared leaves of extant species were prepared following the method of Foster(1952).The terminology used to describe leaf morphology follows Liu (1996) and the Leaf Architecture Working Group(Ellis et al.,2009).
We compared our fossil materials with modernCarpinusspecies and we performed a hierarchical clustering analysis to determine the similarity in leaf morphology between current fossils and modern species.Pictures of extantCarpinuswere referenced from the Chinese Virtual Herbarium(http://www.cvh.ac.cn/),the virtual herbaria of the French National Museum of Natural History(https://science.mnhn.fr/institution/mnhn/item/search/form),the virtual herbaria of the Royal Botanic Garden Edinburgh(https://data.rbge.org.uk/),and an image data set of cleared leaves(Wilf et al.,2021).The hierarchical clustering analysis was performed in PAST.The abbreviation‘HTG’refers to the fossil site,Huatugou town,Qinghai Province,China.
3.Results
3.1.Systematic paleontology
OrderFagales Engler.
FamilyBetulaceae Gray.
GenusCarpinusL.
SectionCarpinusL.
SpeciesC.grandisUnger emend.Heer.
Specimens checked: HTG28001 (Fig.2a and b),HTG28301(Fig.2c),HTG28003 (Fig.2d),HTG28004 (Fig.2e),HTGHTG28308(Fig.2f),and HTG28002 (Fig.2g).
Repository:The Paleontology Laboratory of the School of Earth Sciences,Lanzhou University,China.
Locality: Fossil site HTG028,the Oligocene Shangganchaigou Formation,Huatugou Town,Qinghai Province,China.
Description:Leaves are elliptic,3.5-7.0 cm long by 2.0-3.5 cm wide,the widest part being in the middle;leaf base acute-cuneate(Fig.2a,d,g);leaf apex acuminate without drip tip(Fig.2f,h).Leaf margin is doubly serrate and somewhat irregular,with small teeth,apical side of each tooth convex or concave,and basal side is straight or slightly convex; tooth apices are obtuse,rarely slightly sharp,with acute sinuses between teeth(Fig.2k);about 3-5 teeth present in interval between two consecutive secondary veins.Pinnate primary vein is straight and moderately thick,craspedodromous secondary veins are straight or slightly upwardly curved; secondary veins form an angle of 30-50°with primary vein.In HTG28002 and 28003,11 pairs of secondary veins are preserved,and 8 pairs are preserved in HTG28001,spaced at 0.2-0.8 cm;based on the leaf morphology,leaf apex shape and the space between contiguous two secondary veins(Fig.2),we deduce that the numbers of secondary veins are 12-13.Pectinal veins and external veins are well-developed(Figs.2j and 3a).Close to the leaf margin the secondary veins slightly curved and branched,entering teeth basally and bending sharply upwards,terminating in apices or bases of teeth; branches of secondary veins also entering the nearest teeth(Fig.2l).Basal vein is proximally contiguous with the margin (Fig.2i).Tertiary venation opposite percurrent,tertiaries are almost parallel to each other and nearly perpendicular to the secondary veins (Fig.2j).Higher-order veins are mostly orthogonally reticulate,areoles commonly regular,veinlets are almost absent or simple without branch if they are present (Fig.3e and f).

Fig.3.Comparison of fossil leaves of Carpinus grandis from Huatugou area and extant species of Carpinus and Betula.(a)HTG28001 showing preserved areoles in the dotted region.(b) Betula utilis.(c) C.betulus.(d) C.turczaninowii.In (a),(c) and (d),black arrows denote well-developed external veins.(e) Enlargement of the dotted region in Fig.2f.(f)Enlargement of the dotted region in(a).In(e)-(f),black arrows denote orthogonally reticulate areoles and simple veinlets.(g)Enlargement of the areoles in C.betulus.Black arrows denote orthogonally reticulate areoles and simple veinlets(h)Enlargement of the areoles in B.utilis.Black arrows denote multiple branched veinlets.1° =midvein;2° =secondary vein; 3° = tertiary vein.Scale bars = 10 mm in (a)-(d),1 mm in (e)-(h).
Comparison: Within Betulaceae,the sharp apical bend of the secondary veins on entering the teeth is indicative ofBetula,Carpinus,andOstrya(Wolfe and Wehr,1987),in which this character is present in current specimens(Fig.2l).Compared with these extant genera,Ostryacan be firstly excluded.Manchester and Crane(1987)pointed out that leaves ofOstryaare typically distinguished by secondary veins that are gently curved and/or undulating in course and that have successive prominent abmedial branches to the subsidiary teeth.We examined some clearedOstryaleaves from a recent data set (Wilf et al.,2021) and found that their features support this suggestion,and we found these undulating secondary veins were often present at the base of the leaf.Thereby,Ostryais different from our fossil leaves,which have straight secondary veins.These fossil leaves (Figs.2 and 3) are characterized by an elliptical leaf shape,doubly serrulate leaf margin,craspedodromous secondary veins,percurrent tertiary veins with well-developed areoles,mostly orthogonal reticulate higher-order venation,and the development of free ending veinlets (FEVs); all these features are characteristics of leaves of extant species ofCarpinus(Liu,1996;Worobiec and Szynkiewicz,2007).Some detailed studies have shown that there are two major morphological differences between the leaves ofBetulaandCarpinus: 1) WithinCarpinus,FEVs in areoles are almost absent,or simple if they are present.In contrast,withinBetula,FEVs are well-developed and multiple branched(Wolfe and Wehr,1987; Liu,1996; Worobiec and Szynkiewicz,2007).2) WithinCarpinus,the external veins are well-developed,but they are mostly absent inBetula(Liu,1996).In the current fossil materials,some FEVs can be observed but they are all simple,differing from the multi-branched veinlets inBetula(Fig.3e-h).Additionally,the external veins are well-developed in the current fossils,as those in extantCarpinus(Figs.2d,e,h,and 3a,c,d).In our fieldwork during recent years,noBetulafruits have been found at the site of this fossil leaf(HTG028),and these fruits seem to be only concentrated at the fossil site HTG022.Therefore,we think it is better to assign these fossil leaves toCarpinus.
The Huatugou leaves possess about 12 or 13 pairs of secondary veins,like the species of sectionCarpinus(7-15 pairs),which is different than sectionDistegocarpus(20-34 pairs) (Krüssmann,1976).We compared the leaf morphology of the fossils with modern hornbeams in sectionCarpinus(Table 1).Our comparative analysis indicates that the current fossils are distinguished from most extant species by the acute-cuneate base.In extantCarpinus,species with an acute or cuneate base are rare.A summary of leaf architecture in Betulaceous genera indicated that the most obvious characteristic is thatCarpinus,Corylus,OstryaandOstryopsistypically possess a cordate leaf base(Hickey,1977);an acute or cuneate base,as observed in the current fossil,is more typical ofAlnusandBetula.However,Crane(1981)pointed out that certain leaves with an acute base do occur in species ofCarpinus.We further compared the current fossils with species possessing broadly-cuneate or rounded-cuneate bases in this section.Carpinus monbeigianaHand.-Mazz.,C.tschonoskiiMaxim.,C.polyneuraFranch.,C.mollicomaHu,andC.rupestrisA.Carmus can be excluded because they have more than 14 pairs of secondary veins and setiform teeth.Carpinus omeiensisHu has a broadly-cuneate base and 12-16 pairs of secondary veins,but the serration is simple;C.turczaninowiiHance always has smaller and ovate leaves,different from the current fossils;C.londonianaH.Winkl.andC.kweichowensisHu have cuneate bases but the leaf shape of the former is always lanceolate and the latter have a larger leaf size than our fossils.In the reported fossil species ofCarpinus,those with cuneate bases are rare as well;C.cf.lanceolataHand.-Mazz.from the Miocene Shanwang formation(Shandong,China)has an acute-cuneate base,but the broadly lanceolate leaf shape is different from the current fossils (WGCPC,1978);C.cf.fargesianaH.Winkl.from the late Eocene Lawula formation (Xizang,China) has elliptic leaves and a broadly-cuneate base,but the angle between secondary veins and the primary vein is distinctly unequal on both sides of the primary vein(Tao and Du,1987).Among extant species,the leaf morphology of the Huatugou fossils most closely resembleC.betulusL.(Fig.3),but their tertiary veins are somewhat different.The tertiary veins of our fossil leaves are almost opposite but those ofC.betulusare usually alternate or mixed; in the Huatugou fossils,the number of tertiary veins along the secondaries in the middle of leaves is denser (>15/cm) than inC.betulus(≈10/cm).Among previously reported fossils,our new materials can be assigned to the European fossil speciesC.grandis,which is well known in the Cenozoic floras of Europe and Asia from the middle Oligocene to early Pliocene(Li and Guo,1976;Mai,1981;Tao and Du,1987;Mai and Walther,1988,1991;Hummel,1991;Zastawniak and Walther,1998;Worobiec and Szynkiewicz,2007; Dai et al.,2013).C.grandismost probably is a collective morphotype species and includes several species of fossil leaves; the differences in structure of epidermis in different morphotypes also point to an evolutionary series comprising several species(Worobiec and Szynkiewicz,2007).Leaves ofC.grandisare characterized by the elliptic or oblong-ovate leaf blades and 10-12 pairs of secondary veins,with an acute,obtuse or rounded,rarely cordate base,well-developed and regular areoles,simple veinlets,secondaries slightly curve,tertiary venation percurrent and opposite,tertiaries are nearly perpendicular to the secondary veins,external veins and pectinal veins well-developed,tertiary veins along the secondaries in the middle of leaves dense.All of these characters are similar to the Huatugou fossils.We think it is better to assign the current Oligocene forms into the fossil species rather than to a living species.
Genus:BetulaL.
Subgenus:BetulaL.
Species:Betulasp.
Specimens checked: HTG22031 (Fig.4a),HTG22032 (Fig.4b),HTG22033(Fig.4c and d),and HTG28410 (Fig.4g).
Repository:The Paleontology Laboratory of the School of Earth Sciences,Lanzhou University,China.
Locality: Fossil site HTG022,the Oligocene Shangganchaigou Formation,Huatugou Town,Qinghai Province,China.
Description:Fruits are 2.5-3.0 mm in width and 3.5-5.0 mm in length.Pericarp is ovoid with two thin persistent styles at apex(Fig.4e-g)and two membranous wings on its lateral side.Nutlets are oval or obovate in shape.Membranous wings are 1.6-1.9 mm in width and 2.8-3.3 mm in length; and the width and length of nutlets are 1.4-1.8 mm and 2.3-3 mm respectively.The wings and nutlets are almost equal in width,and the latter are shorter.The widest part of wings is situated at the upper region.
Comparison: The fossil fruits are characterized by two membranous wings,which are as wide as nutlets,or slightly wide.The widest parts of wings are at the upper region.The fruit morphology of these fossils is very similar to those of livingBetulaspecies(Fig.4),although there are some species inAlnusMill.that also have membranous and wide wings.The two genera can be distinguished by the morphology of persistent styles.They are usually slender inBetulabut stout inAlnus(Fig.4i and j).Recent molecular research on the phylogenetic relationship among birch trees divided the genusBetulainto two subgenera,AsperaandBetula,noting that the former has no or very narrow seed wings whereas the latter has obvious seed wings(Wang et al.,2021).Furthermore,the subgenusBetulahas been divided into four sections:Acuminatae,Betula,Costatae,Dahuricae(Wang et al.,2021).Among these,species who have wider wings can be excluded,includingBetula alnoidesBuch.-Ham.ex D.Don andB.luminiferaH.Winkl.It is hard to further identify the current fossil fruits because of poor preservation and a lack of related fossil leaves at this locality.Consequently,we tentatively describe them as an undefined species in subgenusBetula.
4.Discussion
4.1.Nearest living relatives of the Huatugou fossil leaves
We performed hierarchical clustering analysis to determine the similarity in leaf morphology between the fossil leaves and modernCarpinusspecies,based on a data matrix including 33 modern species and 16 leaf characters.Detailed information on the hierarchical clustering is provided in Supplementary data.Before the clustering,we used the principal component analysis (PCA) to assess these characters,gathering the top 12 components with a sum of variance greater than 95%as new variables and obtaining a tree diagram for clustering (for detailed information,see Supplementary data).These analyses indicate that the morphology of the Huatugou fossil leaves most resemblesC.betulus(Fig.5).Thus,we consideredC.betulusas the nearest living analogue of the current fossil leaves.This is consistent with a previous viewpoint(Utescher and Mosbrugger,2015).Unfortunately,associated fruits ofCarpinus,which would help confirm this relationship,have not been found at the Huatugou sites thus far.

Fig.5.Cluster results showing the morphological similarity among current fossil leaves and 33 extant species of Carpinus.For detailed characters and matrix,see Table S1.
4.2.Phytogeographic implications
Recent discoveries revealed a considerably high biodiversity and a humid subtropical ecosystem in central Tibet during the middle Eocene (Liu et al.,2019; Su et al.,2019,2020; Tang et al.,2019).Several Paleogene taxa from this region also revealed a relatively free exchange of lineages across the Northern Hemisphere(Su et al.,2020).For example,Ailanthus maximus(Liu et al.,2019),Cedrelospermum(Jia et al.,2019) andLagokarpos tibetensis(Tang et al.,2019) indicated an obvious floristic linkage between the Tibetan Plateau and European and North American floras in the late Paleogene,a discovery that laid the foundation for exploring the biogeographic significance of the Tibetan Plateau during the early Paleogene.However,more comprehensive information of floristic exchanges between the Tibetan Plateau and other regions in the late Paleogene are still required.
To study the exchange between the current fossil plants and other contemporaneous floras in the Northern Hemisphere,we first gathered the fossil records ofBetulaandCarpinusin the Paleogene(Table 2).Eocene megafossils ofCarpinusare found in northwestern North America,China(Liaoning and Mangkang),and Japan(Fig.6).The earliest unequivocal fruit record ofCarpinuscomes from the middle Eocene (50-49 Ma) of North America (Pigg et al.,2003).Based on fossil records,it is reasonable to assume that the genus dispersed via the Bering Land Bridge,an important corridor for many temperately disjunct lineages until 5.5-5.4 Ma (Milne and Abbott,2002).The earliest reliable fossil ofBetulawas collected from the Paleocene in northeastern China and Kazakhstan(Tao and Xiong,1986).The fossil record ofBetulashows a similar distribution pattern to that ofCarpinus,which likewise suggests an exchange between Northern America and East Asia via the Bering Land Bridge(Fig.6).In addition,the floristic linkage between East Asia and Europe might be weak in the Eocene,given that Turgai Strait still separated Europe and Asia at that time (Mennecart et al.,2018).
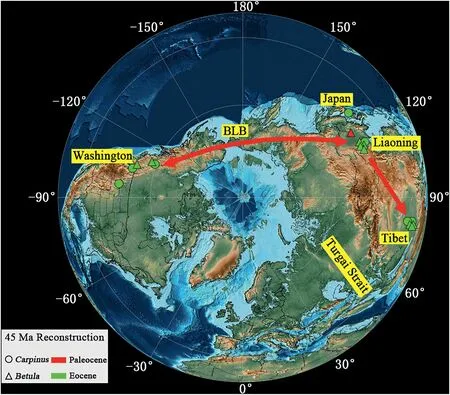
Fig.6.Paleographic map of the world during the early Paleogene (45 Ma).The map was modified from the PALEOMAP PaleoAtlas for Gplates (Scotese,2016).The circles denote fossil records of Carpinus.Triangles denote records of Betula in northwestern North America,Europe,and East Asia.The red line indicates the possible dispersal path.BLB,Bering Land Bridge.
The Betulaceous fossils and other associated fossil angiosperms from the Oligocene of the Qaidam Basin provide further evidence for Paleogene floral interchanges between the Tibetan Plateau and other regions in the Northern Hemisphere.Abundant fossil records indicate thatBetulaandCarpinuswere widely distributed in the Oligocene Eurasia (Fig.7a).As mentioned,the fossil leaves in this study have been assigned toC.grandis,which is a common fossil morphospecies in the Cenozoic deposits of the Northern Hemisphere (Li and Guo,1976; Mai,1981; Tao and Du,1987; Mai and Walther,1988,1991; Hummel,1991; Zastawniak and Walther,1998; Worobiec and Szynkiewicz,2007).In Asia,C.grandishas been found in late Eocene Mangkang and middle Miocene Namling of Tibet (Tao and Du,1987; Guo et al.,2019); and in Europe,this fossil species appears to have had a more extensive distribution since the Oligocene,including the modern-day Czech Republic,southern Russia,and Germany (see Table 2).As the nearest living equivalent of the current fossil leaves,C.betulusis also naturally distributed in central Europe today (Boratynski,2014).We speculate that the current fossilCarpinusfrom the northern Tibetan Plateau might be closely related to European taxa.This speculation is corroborated by the presence of other Oligocene fossil angiosperms found in the Huatugou region.For example,Podocarpium,an extinct genus well-documented in the Cenozoic deposits around Eurasia (Wang et al.,2007; Xu et al.,2015),is abundant in the Oligocene Qaidam Basin (Yan et al.,2018; Han et al.,2020; Song et al.,2020).It is also known from the Oligocene of south China,central Europe,and western Europe (Fig.7a).Recent research has revealed thatPodocarpiummay have originated in East Asia,migrated into the central valley of Tibet in the late Eocene,and then spread westward to Europe (Li et al.,2022).Many fossil samaras and leaves ofAilanthus confuciihave been found at the Huatugou fossil site (Yang et al.,2021a,2021b).This is a species that been discovered in Oligocene deposits from Europe to East Asia.Another fossil species,A.tardensisHably,which is found in Oligocene sediments in Europe and South China,indicates a subsequent diversification in Europe or South Asia(Song et al.,2014).The enhanced floristic affinity between Europe and Asia during the Oligocene can be attributed to the continuous retreat of the Turgai Strait at that time.In addition,enhanced moisture transport via westerlies provided suitable climatic conditions for the exchange of these plants during the Oligocene (Wu et al.,2021).The flora in the Oligocene Huatugou area also shares some components with the contemporaneous floras of Yunnan Province at the southeastern edge of the Tibetan Plateau(Huang et al.,2016;Huang,2017;Tian et al.,2021).With consideration of the higher global temperature(Zachos et al.,2001) and the moderate elevation of the southeastern plateau during the late Eocene to early Oligocene (Li et al.,2015; Spicer et al.,2020; Xiong et al.,2020),the floras in the northeastern and southeastern plateau might have been interconnected at that time.
Overall,the records of plant fossils indicate a close floristic linkage between the Qaidam Basin and Europe during the Oligocene,which can be attributed to the retreat of the Turgai Strait and the favorable climatic conditions provided by the enhanced moisture transport via westerlies.Thus,the Qaidam Basin might represent a pathway on the northern plateau for the free floristic exchange between the Tibetan Plateau and Europe during the late Paleogene.Our findings enrich our knowledge of the biogeographic significance of plant fossils from the Qaidam Basin.Moreover,these fossils reveal the potential influence of westerlies on the formation of paleovegetation in the late Paleogene northern Tibetan Plateau.
AlthoughCarpinusis a common element in the modern North American vegetation,it exhibits an unusual and somewhat perplexing fossil distribution.Reliable records ofCarpinusare extremely rare throughout post-Eocene North American sediments(Manchester,1999).It still remains uncertain ifCarpinuswas present but left no clear fossil record or if the genus went extinct in North America and migrated back to the continent recently(Stults et al.,2002; Stults and Axsmith,2009).After the Paleogene,Carpinuswas still widespread in Europe and East Asia; its modern distribution was mostly shaped since the Neogene.Under the influence of the increasing elevation of the southern Tibetan Plateau and consequent climatic changes (Spicer et al.,2003),this genus was only distributed at low latitudes on and around the plateau(Fig.7b).
4.3.Paleoenvironmental implications
Many Betulaceae species are important components of the deciduous broad-leaf forests in the north temperate zone (Li and Skvortsov,1999).Carpinus betulus,the nearest living equivalent of current fossil leaves,is naturally distributed at low-elevation (not exceeding 2300 m)warm and moist regions in central Europe today(Boratynski,2014).In China,the species of sectionCarpinuslive in similar environments,with mean annual temperature(MAT)values rangingfrom-1.8 to 24.1°C,mean annualprecipitation(MAP)values ranging from 320 mm to 2992 mm,and elevations not exceeding 2800 m (Li and Skvortsov,1999; Fang et al.,2011).Betulaalso commonly inhabits the mid-latitude region in the Northern Hemisphere (Chen,1994b),and some act as keystone species of forests across Eurasia and North America(Ashburner and McAllister,2013).
During our fieldwork in the Huatugou area over the past few years,many other fossil plants were also found from the continuous layer in the lower and middle parts of Shangganchaigou Formation(Yang,2022).DesmanthusWilld.(Leguminosae),collected from lower Shangganchaigou Formation,is a pantropical legume genus native to the New World,revealing that the northwestern Qaidam was much warmer in the early Oligocene (Yang et al.,2021a,2021b).A.confuciiUnger (Simarubaceae),which was found from a nearby layer and is the most common member among fossilAilanthus,is thought to have shared a similar ecological niche with extantA.altissimain the past,also indicating that climate conditions at the fossil site were much warmer and more humid during the early Oligocene than currently(Yang et al.,2021b).In addition,some fossil fruits and leaves from this region,includingPodocarpium(A.Braun) Herendeen,AcerL.,UlmusL.,CupressusL.,ChamaecyparisSpach,ThujaL.,AdiantumL.,andEquisetumL.,have been previously reported(Yan et al.,2018;Chen et al.,2021;Yang,2022).Preliminarily research based on the coexistence approach suggests that the mean annual temperature (MAT) and mean annual precipitation(MAP)was about 14°C and 600-1200 mm,respectively(Yang,2022),which is partly consistent with a recent study (Song et al.,2020) focused on coetaneous northeastern Qaidam Basin(MAT = 11.6 ± 2.4°C,MAP >1000 mm).
4.4.Warm and humid forests in Oligocene Qaidam for floristic exchange
In addition to the fossil plants mentioned above,a paleobotanical study in the western Qaidam Basin noted thatQuercusL.is common in the Shangganchaigou Formation (Zhong,2007).Palynological data from the Dahonggou Section in the northeastern Qaidam Basin also showed an abundance ofQuercusin this formation (Lu et al.,2010).In addition,recent research in the Dahonggou Section indicated that the leaf fossil assemblages from the early Oligocene (30.8 Ma) were dominated byPopulusL.andPodocarpium,with the occasionalCyclocaryaIljinsk (Song et al.,2020).These fossil plants collectively suggest that there used to be a warm and humid deciduous broad-leaf forest during the Oligocene in Qaidam.Few trees today can survive such an elevation at this latitude on the plateau (Fig.1f).The regional eradication of these plants might have resulted from the rising elevation and decreasing temperatures and precipitation since the Miocene (An et al.,2001; Tiffney and Manchester,2001; Garzione et al.,2005).
The global cooling at the Eocene-Oligocene transition (Zachos et al.,2001; Zanazzi et al.,2007) not only profoundly changed the climatic conditions and floristic composition in southeastern Tibet,i.e.,from subtropical/warm temperate in the late Eocene to cool temperate in the early Oligocene (Su et al.,2018),but this climatic event and consequent cooling and aridification (Dupont-Nivet et al.,2007) have also made an impact on the succession of fish fauna in Huatugou (Yang et al.,2018,2022).After the Eocene-Oligocene transition,global temperatures gradually rose(Zachos et al.,2001).In western Qaidam,aridity decreased,which has been attributed to increased moisture from westerlies (Wu et al.,2021).Together,these conditions created a suitable environment in the Qaidam Basin for plants that inhabit warm,humid forest to flourish and eventually spread.
5.Conclusions
In this study,we describe megafossils of Betulaceae collected from the Oligocene western Qaidam Basin (Huatugou area),includingCarpinusleaves andBetulafruits.Our main conclusions are as follows:(1)Based on detailed morphological comparison,the fossil leaves were assigned toC.grandisand the fossil fruits were identified asBetulasp.They represent the first fossil record of Betulaceae in the Qaidam Basin.(2) These Betulaceous fossils,together with previously reported fossil plants from the same locality,reveal a close floristic linkage between the Qaidam Basin and Europe during the Oligocene.The Qaidam Basin might represent a pathway on the northern plateau for the free floristic exchange between the Tibetan Plateau and Europe during the late Paleogene.(3)Fossil plants collected from Huatugou area suggest a warm and humid deciduous broad-leaf forest during the Oligocene.The westerlies provided suitable climatic conditions for such a warm and humid forest,allowing the Qaidam Basin to act as a region for these plants to flourish and spread.
Author contributions
D.Y.and T.Y.designed research and led the expeditions;J.C.,H.C.,L.H.,L.Z.,W.L.,J.W.,and X.L.collected the fossil and extant specimens;Y.D.,W.L.,and S.X.performed research;T.Y.and J.C.analyzed data; and D.Y.and T.Y.wrote the paper.
Declaration of competing interest
The authors declare they have no conflicts of interest.
Acknowledgements
We thank to Dr.Junling Dong (Chengdu University of Technology,Chengdu),and Dr.Pengju He(Lanzhou University,Lanzhou)for substantial suggestions; Dr.Haylin Chan and Pallas Cate (Sun Yatsen University,Guangzhou) for helpful remarks and assisting with English; Prof.Steven R.Manchester (University of Florida,U.S.A.) for providing helpful suggestions and literature; Dr.Jianwu Li,Mr.Bing Wang (Xishuangbanna Tropical Botanical Garden,Mengla),and Prof.Trevor Hodkinson (Trinity College,Ireland) for providing some extant specimens ofCarpinusandBetula.We sincerely thank Dr.Bian Wang (IVPP,Beijing) for improving the language.This research was conducted under the China Postdoctoral Science Foundation (No.2022M723151); the Second Tibetan Plateau Scientific Expedition Research Program (No.2019QZKK0704);the National Natural Science Foundation of China(No.42172005,41272026,41972008,31870200);and the Strategic Priority Research Program (B) of the Chinese Academy of Sciences(XDB26000000).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2023.03.007.

- 植物多樣性的其它文章
- Flower morphology of Allium (Amaryllidaceae) and its systematic significance
- Phylotranscriptomic discordance is best explained by incomplete lineage sorting within Allium subgenus Cyathophora and thus hemiplasy accounts for interspecific trait transition
- How to fill the biodiversity data gap:Is it better to invest in fieldwork or curation?
- Life forms affect beta-diversity patterns of larch forests in China
- A phylogenetic approach identifies patterns of beta diversity and floristic subregions of the Qinghai-Tibet Plateau
- Community structure and species diversity dynamics of a subtropical evergreen broad-leaved forest in China: 2005 to 2020