
Benthodytes occidentpalauta sp.nov., a new species of deep-sea holothuroid (Elasipodida: Psychropotidae) from the west of Kyushu-Palau Ridge in the Western Pacific Ocean*
2024-02-27 08:28ChongzhenYUANChunshengWANGDongshengZHANG
Chongzhen YUAN, Chunsheng WANG,2,3,**, Dongsheng ZHANG,2,3
1 Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China
2 Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519082, China
3 School of Oceanography, Shanghai Jiao Tong University, Shanghai 200030, China
Abstract Benthodytes occidentpalauta sp.nov.was collected from the Kyushu-Palau Ridge at a depth of 5 481 m in 2021.This new species is characterized by a gelatinous body wall, violet skin, six pairs of dorsal papillae, and a rough mid-ventral surface without tube feet.The dorsal deposits are rod-shaped and tripartite.Two types of papillae deposits as crosses with four arms with central bipartite apophyses.Ventral deposits are rods.Tentacle ossicles are rod-shaped with end protrusions.Gonad deposits are rodshaped, tripartite, and cross-shaped.The phylogenetic analyses based on cytochrome oxidase subunit 1(COI) and 16S individually and a concatenated dataset of COI and 16S genes of this species support that B.occidentpalauta sp.nov.belongs to Benthodytes.
Keyword: Holothuroidea; Benthodytes; cytochrome oxidase subunit 1 (COI); 16S; phylogenetic analysis
1 INTRODUCTION
Holothurians (Echinodermata, Holothuroidea)are a group of animals in the phylum Echinodermata.The number of reported species of holothurians exceeds 1 779 (Paulay and Rogacheva,2022).Limited by sampling, holothurians in the deep sea are relatively difficult to study compared to those in offshore and shallow waters.However,holothurians are crucial for the research of deep-sea benthic organisms.In some deep-sea trenches,holothurians constitute more than 90% of the biomass (Belyaev, 1972).
The order Elasipodida, family Psychropotidae and genusBenthodyteswere established by Théel in the scientific expedition report of H.M.S.“Challenger” in 1873-1876 (Théel, 1882).Benthodytesis defined by Théel as a group with ventral mouth, posterior anus, more or less depressed body with large anterior brim, twelve to twenty tentacles.The tube feet ofBenthodytesare arranged in a single row around the brim of the body and in a double row along the odd ambulacrum.Benthodytescurrently includes 15 valid species that have been recorded (WoRMS, 2022):Benthodytes abyssicolaThéel, 1882;BenthodytessanguinolentaThéel, 1882;BenthodytestypicaThéel, 1882;BenthodytesincertaLudwig, 1893;Benthodytes linguaPerrier R., 1896;BenthodytessibogaeSluiter,1901;BenthodytessuperbaKoehler & Vaney, 1905;BenthodytesplanaHansen, 1975;Benthodytes valdiviaeHansen, 1975;BenthodytesgosarsiGebruk, 2008;BenthodyteswolffiRogacheva &Cross in Rogacheva et al., 2009;BenthodytesvioletaMartinez, Solis-Marin & Penchaszadeh, 2014;BenthodytesmanusensisXiao, Li & Sha, 2018;BenthodytesmarianensisLi, Xiao, Zhang & Zhang,2018; andBenthodytesjiaolongiYu, Zhang, Zhang& Wang, 2022.On the basis of phylogenetic studies,Benthodyteswas regarded as a paraphyletic group including two clades (Yu et al., 2022).One clade includedB.sanguinolentaandB.jiaolongiand the other clade containedB.marianensis,B.manusensis,B.palauta, andB.sibogae.
Some holothurian specimens were collected in the Kyushu-Palau Ridge during the cruise of DY68 in November 2021.According to the database of Ocean Biodiversity Information System (OBIS)from 1906 to 2018, eight species of psychropotids have been reported adjacent to Kyushu-Palau Ridge,includingPsychropotesverrucosaLudwig, 1893,PsychropotesraripesLudwig, 1893,Psychropotes longocaudaThéel, 1882,PsychropotesdepressaThéel, 1882,B.typica,B.sanguinolenta,B.lingua,andB.incerta(https://obis.org/taxon/123194).In addition, five species of psychropotids, identified asB.marianensis,B.manusensis,B.palauta,B.jiaolongi, andPsychropotesverrucicaudatusXiao, Gong, Kou & Li, 2019, have recently been reported in this area (Li et al., 2018; Xiao et al.,2018, 2019; Yu et al., 2021, 2022).
Based on the morphological and phylogenetic results,Benthodytesoccidentpalautasp.nov., a new species of Psychropotidae, was identified as new to science.The results of the phylogeny support theB.occidentpalautasp.nov.belonging to the clade containingB.marianensis,B.manusensis,B.palauta,andB.sibogae.
2 MATERIAL AND METHOD
2.1 Sampling and morphological observation
The sample (RSIO6820405) was collected during the cruise of DY68 at a depth of 5 481 m from the Kyushu-Palau Ridge (129.933°E, 16.796°N) by theJiaolongHuman Operated Vehicle (HOV) on November 23, 2021.Before preservation, a Canon EOS 7D camera (Canon Inc., Tokyo, Japan) was used to take photographs of the specimen.A tissue sample was then cut and stored in a -80-°C refrigerator for DNA extraction.Finally, the samples were fixed in 4% seawater-formaldehyde solution and stored in the Repository of the Second Institute of Oceanography (RSIO).Sodium hypochlorite(10%) was used to dissolve body tissues (dorsum,papillae, ventrum, brim, tentacles, and gonad) for extraction of deposits, which were sequentially washed and dehydrated with 80%, 90%, and 100%alcohol.Deposits were dried for one day and observed using a scanning electron microscope (TM 1000; Hitachi, Ltd., Tokyo, Japan).
2.2 PCR amplification and phylogenetic analysis
Total genomic DNA was extracted from 25 mg of muscle tissue using a DNeasy Blood & Tissue Kit(QIAGEN, Hilden, Germany) according to the manufacturer’s instructions.The primers COI-ef/er and 16S-arL/brH (Miller et al., 2017) were used to amplify partial sequences of cytochrome oxidase subunit 1 (COI) and 16S rRNA.The PCRs were performed using a 25-μL reagent mix containing 12.5-μL 2×Phanta Max Master Mix (Vazyme,Biotech Co., Ltd., Nanjing, China), 8.5-μL doubledistilled water, 1 μL of each primer, and 1-μL template DNA, as suggested by the manufacturer.The PCR amplification procedure of COI was as follows: denaturation at 95 °C for 3 min, followed by 40 cycles of 95 °C for 40 s, 45 °C for 40 s, and 72 °C for 50 s, and a final annealing at 72 °C for 5 min.The PCR amplification procedure of 16S was as follows: denaturation at 95 °C for 3 min,followed by 35 cycles of 95 °C for 40 s, 50 °C for 40 s, 68 °C for 50 s, and a final annealing at 68 °C for 5 min.PCR products were confirmed by 1%agarose gel electrophoresis.PCR products with clear electrophoretic bands were sequenced on an ABI 3730xl DNA analyzer (Sangon Biotech Co.,Ltd., Shanghai, China).
Some relevant COI and 16S gene sequences were retrieved from NCBI and downloaded for phylogenetic analysis in this study (Table 1).Two species of the genusPannychiawere used to root the tree.All sequences were managed with PhyloSuite v1.2.2 (Zhang et al., 2020).COI and 16S sequences were aligned with MAFFT (Katoh and Standley, 2013) using the E-INS-I strategy.Ambiguously aligned fragments were removed using Gblocks (Talavera and Castresana, 2007) with the following parameter settings: minimum number of sequences for a conserved/flank position (8/8),maximum number of contiguous nonconserved positions (8), minimum length of a block (2), and allowed gap positions (with half).PhyloSuite(Zhang et al., 2020) was used to concatenate genes into the dataset (COI/16S=638/523 bp).ModelFinder(Kalyaanamoorthy et al., 2017) was used to select the best-fit partition model (edge-linked) using the Akaike information criterion (AIC).Best-fit model according to AIC: GTR+F+I+G4: COI; GTR+F+I: 16S.Maximum likelihood phylogenies were inferred using IQ-TREE (Nguyen et al., 2015) under an edge-linked partition model for 1 000 standard bootstraps.Bayesian inference phylogenies were inferred using MrBayes 3.2.6 (Ronquist et al., 2012)under a partition model (2 parallel runs, 2 000 000 generations), in which the initial 25% of sampled data were discarded as burn-in.MEGA11 (Tamura et al., 2021) was used to calculate pairwise distance for COI and 16S between sequences.Analyses were conducted using the Kimura 2-parameter model(Kimura, 1980).All ambiguous positions were removed for each sequence pair (pairwise deletion option).

Table 1 Gene sequences of the specimens used in this study
3 RESULT
3.1 Systematics and type material examined
Class Holothuroidea Blainville, 1834
Order Elasipodida Théel, 1882
Family Psychropotidae Théel, 1882
GenusBenthodytesThéel, 1882
Benthodytesoccidentpalautasp.nov.(Figs.1-4)
Holotype
One adult specimen, catalog number RSIO6820405, GenBank accession numbers OP538683 for 16S and OP538553 for COI,collected on November 23, 2021 from a sea area(named unofficially as “KPR6”) located in the west of Kyushu-Palau Ridge (129.933°E, 16.796°N) at a depth of 5 481 m.
Type locality
The KPR6 sea area is located on the Kyushu-Palau Ridge, tropical Western Pacific Ocean, at a depth of 5 481 m.
Etymology
From the Latinoccidentalismeaning west.The species name refers to the type locality west of the Kyushu-Palau Ridge.
Diagnosis
Gelatinous body wall, with an extended, slightly swollen cylindrical body.Mouth ventral and anus dorsal.Wider at the head and slightly narrowed at the end.The body dark purple in situ.Brim well developed but constricted.Six pairs of papillae are in two rows along the each dorsal radius.A few processes between the first and second pairs of papillae.Mid-ventral tube feet lacking.The ventral surface dark-colored and rugose.Brim with tips of enclosed tube feet constricted and darker than the surrounding.Thirteen shield-shaped tentacles.Circum-oral papillae present but not visible after preservation in formaldehyde-seawater.The internal organs include one drop-shaped 39-mm-long Polian vesicle with one basal dichotomous gonad.Respiratory tree absent.Calcareous ring are not visible.The dorsal deposits rod-shaped and tripartite.Papillae deposits as crosses with three or four arms with a central bipartite apophysis, and rod-shaped.Rods present in the ventral surface and tentacle.Gonad deposits rod-shaped, tripartite and cross-shaped.
Description of the holotype

Fig.1 Benthodytes occidentpalauta sp.nov.(Holotype,RSIO6820405)
Photographs taken in situ show the sample as a slightly swollen, extended cylinder (Fig.1).The head is disc-shaped with the mouth at the center.The circum-oral papillae are obvious (Fig.1c).Slightly narrowed posteriorly, with well-developed and constricted brim.Tips of enclosed tube feet present on the brim.The full length before preservation with 4% seawater-formaldehyde was approximately 26.9 cm, and the width was approximately 7.3 cm (Fig.1b-c).After preservation,the length was approximately 15.4 cm, and the width was approximately 5.0 cm (Fig.2a-b).The mouth is located on the ventral side, and the anus is located on the dorsal side.The body wall is loosely gelatinous.The specimen in situ was dark purple in color, purplish red on the deck before preservation,and deep purple after preservation with 4%seawater-formaldehyde.After preservation, the color of the area around the mouth is the darkest.The color of the mid-ventral surface is deep but not as deep as the area around the mouth.The other parts of the body wall are the lightest and whitish,mid-ventral tube feet are lacking.The ambulacrum appears as a folded band that extends from below the mouth to the end of the body (Fig.2b).The texture of the ambulacrum is stiffer than that of the surrounding body wall.The tentacles are plump and shield-shaped, with a hard texture.The number of tentacles is 13.The circum-oral papillae are obvious on the deck.However, they are not visible after preservation with seawater-formaldehyde.The right dorsal papillae of the specimen are partially missing.The following description assumes that the left and right dorsal papillae are symmetrical.There are six pairs of papillae on the dorsum.The papillae are constricted after preservation and appear as dark purple patches.The first, second and third pairs of papillae are distributed in the upper part of the body wall.The distance between the first and second pairs of papillae is longer than the distance between the second and third pairs of papillae.There are some small purple spots between the first and second pairs of papillae.These spots are probably the traces of small papillae.The fourth, fifth and sixth pairs of papillae are distributed in the lower part of the body wall.The distance between the fourth and fifth pairs of papillae is shorter than the distance between the fifth and sixth pairs of papillae.The sixth pair of papillae was not at the same distance from the anus,with the left papillae being closer to the anus.

Fig.2 Drawing of Benthodytes occidentpalauta sp.nov.(Holotype, RSIO6820405)
The dorsal body wall deposits are rod-shaped of three types and tripartite.The thick rod-shaped deposits (Fig.3a) are approximately 600 μm long and approximately 45 μm wide at the widest point.The slender rod-shaped deposits (Fig.3b) are approximately 560 μm long and 30 μm wide.The rod-shaped deposits with 31-μm middle processes(Fig.3c) are approximately 570 μm long and 30 μm wide.The arms of tripartite deposits (Fig.3d) are 120-170 μm in length.

Fig.3 Scanning electron microscope images of deposits from Benthodytes occidentpalauta sp.nov.(Holotype,RSIO6820405)
Papillae deposits two types of crosses with four arms and central bipartite apophyses, two types of rods and one type of tripartite.The smaller type of crosses (Figs.3e & 4a), in which arms bend at arm 1/2, are 115 μm in length.There are some obvious spines on 1/2 of the central apophyses near the base.The whole central apophysis is approximately 190 μm long.The bipartite arms of central apophyses are 60-70 μm in length with nearly horizontal arms at an angle of approximately 170°with 5-6 vertical downward spines per arm.The larger type of crosses (Figs.3f-g & 4b), in which the arms are straight with spines, are 265 μm in length.The whole central apophysis is approximately 100 μm long.The bipartite arms of central apophyses are 200 μm in length with approximately 4 spines per arm.There are two types of rodshaped deposits.The thick rod-shaped deposits are approximately 695 μm long and 48 μm wide at the widest part with small spines at the end (Fig.3h).The slender rod-shaped deposits are 320-570 μm in length and 15-25 μm in width (Fig.3i-k).The long arms of tripartite deposits are 190 μm in length,while the short arm is 90 μm in length.

Fig.4 Drawing of deposits from Benthodytes occidentpalauta sp.nov.
Two types of ventral surface deposits as rods.The thicker type (Fig.3l-m) is approximately 560-700 μm long and 40-50 μm wide, with a few spines at the ends.The slender type (Fig.3n-o) is approximately 320-660 μm long and 15-30 μm wide, with a few spines at the ends.
Two types of tentacle deposits as rods.The larger type (Fig.3p-s) is approximately 820-1 200 μm long and 80-95 μm wide, with a flat middle without spines.Spines present at the end.Some are terminally bipartite at one side (Fig.3s).The smaller type (Fig.3t-v) is approximately 520-580 μm in length and 40-65 μm in width, with 2-4 spines at the end.
The gonad deposits (Fig.3w-y) rod-shaped,tripartite and simple cross-shaped without apophysis.The rods are 600-670 μm in length and approximately 15 μm in width.Spines occasionally occur on the rods.The long arms of tripartite deposits are 660-700 μm in length, while the short arm is 140-150 μm in length.Bifurcation is present on the end of the short arm for some tripartite deposits.The long arms of crosses are 270-350 μm in length, while the short arm is approximately 145 μm in length.Spines occur occasionally on the arms.
Relationship
Benthodytesoccidentpalautais similar toB.marianensisandB.incerta.The characteristics ofB.marianensisare summarized as follows (Li et al.,2018): nine pairs of dorsal papillae.Other papillae of the same or smaller size distributed on the front of the body.Tentacles, at least 12, retracted into stalks.Information on mid-ventral tube feet is lacking.Tentacle deposits are large rods.Dorsal deposits are crosses with four arms and central bipartite, tripartite or tetrapartite apophyses.There is also a type of special crosses with four arms and high apophyses that end in two horizontal branches.Ventral deposits are rod-shaped with central bipartite apophyses and crosses of three or four arms with central rudimentary apophyses.The characteristics ofB.incertaare summarized as follows (Hansen, 1975): 6-9 dorsal papillae along each radius.Mid-ventral tube feet forming an irregular double row.Tentacles 15.Papillae deposits as large crosses with four arms and a central bipartite apophysis and small crosses with four arms and a central bipartite apophysis that end in two horizontal branches.Ventral deposits usually reduced;apophyses, when present, undivided.
The differences between the new species andB.marianensisare summarized as follows.(1) The brim of the new species is not as broad as the brim ofB.marianensis.(2) The number of papillae of the new species is lower than that ofB.marianensis(6 pairs in the new species and 9 pairs inB.marianensis).The dorsal surface of the new species is smoother than that ofB.marianensiswithout any other small papillae.(3) The papillae deposits of the new species are rods, crosses with three arms and crosses with four arms and a central bipartite apophysis, while the papillae deposits ofB.marianensisare crosses with four arms and bipartite, tripartite or tetrapartite apophyses.The differences between the new species andB.incertaare summarized as follows.(1) The ambulacrum of the new species is rougher without tube feet.The tube feet ofB.incertaform an irregular double row.(2) The sizes of crosses with four arms and a central bipartite apophysis with horizontal branches are different.The arms of the new species are longer than the arms ofB.incerta(115 μm for the new species, 70 μm forB.incerta).Moreover, barbed spines are present on the central apophysis of new species.The central apophysis ofB.incertais smooth.(3) The sizes of crosses with four arms and a central bipartite apophysis with angled branches are different.The arms of the new species are shorter than the arms ofB.incerta(265 μm for the new species, 750 μm forB.incerta).(4) In addition to the dorsal papillae, rod-shaped deposits are present in the dorsum, ventrum, tentacles, and gonads of new species.Sparse numbers of rods and crosses with four arms were present in the ventrum ofB.incerta.(5) No primitive crosses with four arms present in any part of the new species but present in the ventrum ofB.incerta.Crosses with three arms present in the dorsum of the new species but not present inB.incerta.
The appearance ofB.occidentpalautasp.nov.is consistent withBenthodytesand differs fromB.marianensis,B.incertaand any other species inBenthodytes.Thus, it can be recognized as a new species.
Distribution
Only in the type locality.
3.2 Molecular phylogenetic analyses

Fig.5 Phylogenetic trees of family Psychropotidae based on a concatenated dataset of COI and 16S genes
The molecular phylogenetic trees (ML and BI)reconstructed based on the COI-16S data are shown in Fig.5.The trees based on the COI gene are shown in Fig.6.The trees based on the 16S gene are shown in Fig.7.All trees showed that the genusBenthodyteswas not monophyletic.GeneraPsychropotesandPsycheotrepheswere nested within the genusBenthodytes.This indicates that the classification of the genusBenthodytesshould be reconsidered.In the COI-16S tree and COI tree,B.occidentpalautasp.nov.clustered together withB.marianensis,Benthodytessp.SB-1-11,B.manusensis,B.sibogae, andB.palauta, forming a well-supported clade (COI-16S tree: BPP=1,BS=100; COI tree: BPP=1, BS=93).Benthodytes occidentpalautasp.nov.was located in the lateral clade of the sister group.In the 16S tree,B.occidentpalautasp.nov.clustered withB.marianensis, forming a low-supported clade (BPP=0.616, BS=60).This suggests that the topology of the 16S tree is not suitable for supporting the evolutionary relationships of the new species.All phylogenetic trees support the separation ofB.occidentpalautasp.nov.from other congeners.
The divergence of COI and 16S are shown in Tables 2 & 3.The result shows thatB.occidentpalautais closely related toB.marianensis(COI: 0.062 6;16S: 0.013 5).This supports the results of morphological analysis.In addition, for the clade thatB.occidentpalautalocated in, the maximum interspecific divergence distance of the COI is 0.098 1 (betweenB.occidentpalautaandB.palauta).And the maximum interspecific divergence of 16S is 0.038 6 (betweenB.occidentpalautaandB.palauta).Moreover, the minimum COI divergence betweenB.jiaolongiand the species of the clade thatB.occidentpalautalocated in is 0.206 7 (betweenB.jiaolongiandB.marianensis), which is 2.1 times of the maximum interspecific divergence described above.As for 16S, the minimum pairwise distance is 0.192 6 (betweenB.jiaolongiandB.palauta),which is 5.0 times of the maximum interspecific divergence described above.ForB.sanguinolenta,the minimum COI divergence is 0.187 3 (betweenB.sanguinolentaandB.occidentpalauta), which is 1.9 times of the maximum interspecific divergence described above.This supports the results of the evolutionary tree analysis that taxonomic status within the genusBenthodytesshould be reconsidered.
4 DISCUSSION
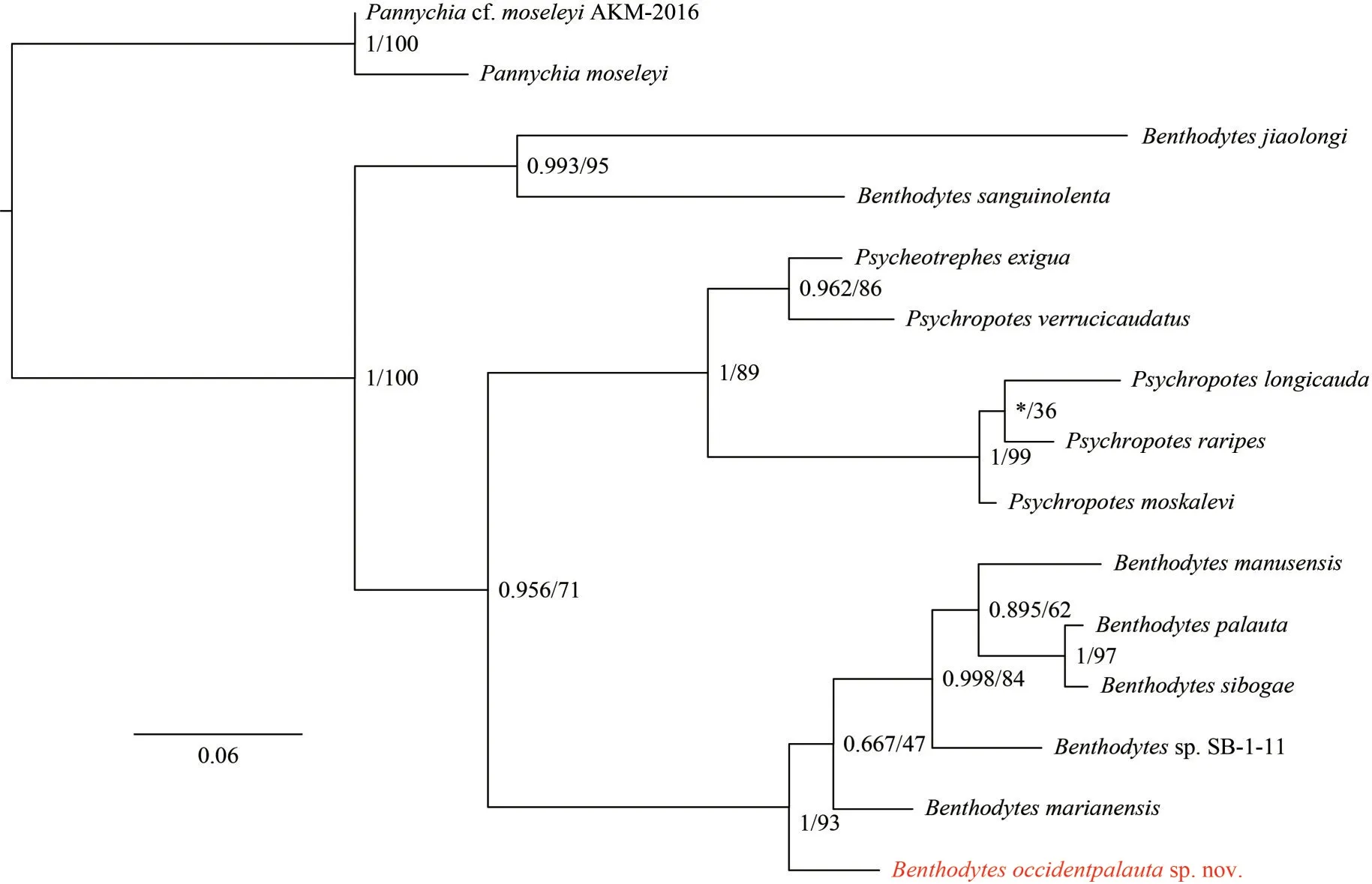
Fig.6 Phylogenetic trees of family Psychropotidae based on a concatenated dataset of COI genes

Fig.7 Phylogenetic trees of family Psychropotidae based on a concatenated dataset of 16S genes
Benthodytesoccidentpalautasp.nov.is morphologically distinguished from the other species ofBenthodytesby the rough ambulacrum without the tube feet, the dark tips of enclosed tube feet at the brim, the six pairs of dorsal papillae, the two types of crosses with four arms and central bipartite apophyses of papillae, the rod-shaped and tripartite dorsal deposits, and the rod-shaped deposits from the ventral surface, tentacles and gonads.The absence of ventral tube feet does not suggest natural abrasion; if it did, there would be at least some tracesof tube feet.Therefore, this should be a characteristic of this new species.Based on morphological characteristics,B.occidentpalautasp.nov.is closely related toB.incertaandB.marianensis.They shared similar body shapes and similar crosses, especially a type of special crosses with four arms and central bipartite apophyses that end in two horizontal branches.However, the new species differs fromB.marianensisin the number and position of dorsal papillae.The new species differs fromB.incertaby the lack of tube feet.Regarding deposits, the new species differs fromB.marianensisandB.incertaby the rod-shaped deposits in the dorsum and ventrum.In this study, we also described the deposits from the tentacle and gonad.The rodshaped deposits of tentacles are much larger than dorsum rods and ventrum rods (820-1 200 μm long for tentacles, 695 μm long for papillae, 700 μm long for ventrum, 596 μm long for dorsum).

Table 2 Estimates of COI divergence between sequences

Table 3 Estimates of 16S divergence between sequences
Based on the morphological characteristics, the species inBenthodytesfall into two distinct groups of relationships according to Hansen, 1975(Hansen, 1975): (1)B.incerta,B.lingua,B.valdiviae,B.sibogae, andB.planahave regular cross-shaped deposits with a large bipartite central apophysis and 15 or fewer tentacles.(2)B.typicaandB.sanguinolentahave strongly reduced, rod-shaped deposits and a high tentacle number (15-20 inB.typicaand 18 inB.sanguinolenta).
Benthodytesabyssicola,B.gosarsi,B.manusensis,B.marianensis,B.palauta, andB.occidentpalautasp.nov.have regular cross-shaped deposits with a large bipartite central apophysis and a low tentacle number(15 inB.abyssicola, 10 inBgosarsi, 12 inB.manusensis, at least 12 inB.marianensis, 10 inB.palauta, and 13 inB.occidentpalauta) as first group (Gebruk, 2008; Rogacheva et al., 2009; Li et al.,2018; Xiao et al., 2018; Yu et al., 2021).These species should be considered the first group.B.superbabelongs to the first group according to Hansen(1975) given the irregular, bipartite apophyses without crosses and an unknown number of tentacles.The phylogenetic results supportB.marianensis,B.manusensis,B.palauta,B.sibogae,B.occidentpalautasp.nov.cluster into one clade.B.occidentpalautasp.nov.is located in the lateral clade of the first group.
BenthodytesjiaolongiandB.wolffihave reduced,rod-shaped deposits and a high tentacle number (18 inB.jiaolongiand 20 inB.wolffi) as the second group (Rogacheva et al., 2009; Yu et al., 2022).The phylogenetic results support that they should be considered the second group.In addition, according to the results of the phylogenetic tree, the interspecific genetic distance of this clade is very long.The divergence of COI betweenB.jiaolongiandB.sanguinolentais 0.191 3.A study by Uthicke et al.(2010) reported that, for COI, the average congeneric divergence of holothurians is 16.9%.Byrne et al.(2010) reported that sequence divergence between species within the genusStichopusis 1.1%to 16.2% for COI and 1.4% to 15.3% for 16S.It is clear that the divergence between the clades of genusBenthodytesexceeds that of the preceding congeneric divergence for COI and 16S.This supports that taxonomic status within the genusBenthodytesshould be reconsidered.
5 CONCLUSION
Morphological analyses identifiedBenthodytes occidentpalautasp.nov.as a new species.Phylogenetic analyses revealed the separation ofB.occidentpalautasp.nov.from congeners of the genusBenthodytes.The new species is characterized by mid-ventral ambulacrum without tube feet, rodshaped deposits in the dorsum and ventrum, and papillae crosses with four arms and high apophyses that end in two horizontal branches.B.occidentpalautasp.nov.is the species of Elasipodida reported west of the Kyushu-Palau Ridge.The information on this new species will be useful for the taxonomic study of the genusBenthodytesand the diversity of psychropotid species in the area of Kyushu-Palau Ridge.
6 DATA AVAILABILITY STATEMENT
The data generated and analyzed to support the research result can be provided by the corresponding author upon reasonable request.
7 ACKNOWLEDGMENT
The authors are grateful to the assistance of the crew ofJiaolongHOV with sample collection and in-situ photographs.Thanks to Bo LU and Chengcheng SHEN from the Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, for taking the photos and preserving the samples on deck.We are grateful to the reviewers for their constructive comments.
 Journal of Oceanology and Limnology2024年1期
Journal of Oceanology and Limnology2024年1期
- Journal of Oceanology and Limnology的其它文章
- Contrasts of bimodal tropical instability waves (TIWs)-induced wind stress perturbations in the Pacific Ocean among observations, ocean models, and coupled climate models*
- Variability of the Pacific subtropical cells under global warming in CMIP6 models*
- Identification of thermal front dynamics in the northern Malacca Strait using ROMS 3D-model*
- Magmatic-tectonic response of the South China Craton to the Paleo-Pacific subduction during the Triassic: a new viewpoint based on Well NK-1*
- An improved positioning model of deep-seafloor datum point at large incidence angle*
- Microplastics in sediment of the Three Gorges Reservoir:abundance and characteristics under different environmental conditions*